
N.C. Anusha
Sime Darby Technology Centre, UPM-MTDC III, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
M.S. Umikalsom
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
T.C. Ling
Department of Food Process Engineering, Faculty of Engineering, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
A.B. Ariff
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
Asian Journal of Biotechnology
Year: 2012 | Volume: 4 | Issue: 1 | Page No.: 15-29







ABSTRACT
Relationship between fungal growth morphology and the ability to produce lipase in solid state fermentation was studied employing four fungal strains (A. niger DSMZ 2466, A. niger SDTC SRW-4, T. viride SDTC EDF 002 and A. terreus DSMZ 5770). The fermentations were performed in 500 mL flask using palm kernel cake as substrate. Scanning Electron Microscope (SEM) was used to examine growth morphology of the fungi. Pearson correlation and regression analyses performed on quantified morphological dimensions and the fungal growth, enzyme productivity and total protein were able to generate predicted values of equation models. All four strains showed good growth on palm kernel cake at day 6 of fermentation ranging from 0.49 to 0.71 mg g-1 dry substrate. Among these strains, substantial lipase activity was only obtained by A. niger DSMZ 2466 and A. niger SDTC SRW-4 which also showed positive relationship between spore diameter (quadratic, R2; 0.999) and branch diameter (linear, R2; 0.971) with lipase activity. T. viride SDTC EDF 002 and A. terreus DSMZ 5770 showed positive correlations for tip length and tip diameter and size of hyphae diameter and the total protein content in the fermentation substrate. Results from this study showed that the fungi with larger spore diameter and branch diameter contributed to higher ability in producing lipase.
PDF Abstract XML References Citation
Received: October 31, 2011;
Accepted: January 07, 2012;
Published: February 23, 2012
How to cite this article
N.C. Anusha, M.S. Umikalsom, T.C. Ling and A.B. Ariff, 2012. Relationship between Fungal Growth Morphologies and Ability to Secrete Lipase in Solid State Fermentation. Asian Journal of Biotechnology, 4: 15-29.
DOI: 10.3923/ajbkr.2012.15.29
URL: https://scialert.net/abstract/?doi=ajbkr.2012.15.29
DOI: 10.3923/ajbkr.2012.15.29
URL: https://scialert.net/abstract/?doi=ajbkr.2012.15.29
INTRODUCTION
Lipase enzymes have achieved a prominent positioning in the global market with continuous increase in market demand. The huge and attractive industrial applications of lipases have moved this enzyme into the third group in volume of sales after proteases and amylases (Pandey, 2003; Mahadik et al., 2002; Freire et al., 1997). Lipases are able to produce by various microorganisms, however, fungi which is widely known as one of the best lipase sources can be easily cultured in Solid State Fermentation (SSF) where growth and secretion of the product may occurs on the surface of the solid support and within the support matrix.
Ample of research has been conducted to enhance lipase production by fungi in SSF. The parameters that were evaluated include supplementation of specific nutrients to the solid substrate, pH, temperature, moisture content and aeration. Moreover, in the design and operation of bioreactor employing fungus, knowledge on fungal morphology is one of the key factors need to be considered. In SF, variation in hydrodynamic conditions may influence growth morphology of the fungi. On the other hand, growth is restricted to the surface of the solid matrix in SSF, where details structural information of growth morphology is difficult to be studied. Reports on the effect of fungal growth morphology on the performance of SF are available in the literature (Teng et al., 2009; Haack et al., 2006; Spohr et al., 1997; Oncu et al., 2007; Lim et al., 2006), however, reports concerning morphological aspects of the fungi and their relationship with metabolites production in SSF are limited. However, at the microscopic level, there is evidence to suggest that extracellular protein yield is positively correlated with increased numbers of actively growing hyphal tips (Wessels, 1993; Gordon et al., 2000; Muller et al., 2002). It has also been suggested that more branching leads to more tips which should result to more products (Wosten et al., 1991).
In fermentation employing fungus, productivity can be increased if the growth morphologies could be controlled at the required characteristics (Ahamed and Vermette, 2009). Moreover, Teng et al. (2009) has described in the research paper that enzymes, primary and secondary metabolites has been affected by the fungal morphology. Growth morphology of the fungi can be divided into two types, (1) micromorphology and (2) macromorphology. Micromorphology has direct influences on enzyme biosynthesis and secretion capacity. On the other hand, macromorphology influences rheology and the mass transfer properties of the culture. Micromorphology deals with the morphology of the individual hyphal elements such as diameter and length of the hyphal elements as well as number of tips of an individual hyphal (Teng et al., 2009).
Fungal morphology has been established in several processes to lead to more productive industrial processes, however, to-date a clear relationship between the morphology and the product enhancement has not been well established as described by Grimm et al. (2005). This is due to the difficulties into investigating many interrelated factors that may affect both the fungal morphology and the product formation. Hwang et al. (2004) has suggested that quantitative structural information of the fungal morphology would need to be experimented to obtain a complete understanding of this relationship.
The objective of this study was to evaluate the relationship between the micro-morphology characteristics and growth kinetics of several fungi species on extracellular lipase production in SSF. Several types of lipase producing-fungi that have potential for industrial processes were used as the model systems. Micromorphology dimensions of the fungi which include spore diameter, hyphae diameter, branch diameter, tip diameter and tip length and their relationship with the ability to produce lipase was studied. The information generated from this study may give better understanding on fungal growth kinetics and their relationship to micro-morphological forms and productivity in industrial SSF.
MATERIALS AND METHODS
Microorganisms and inoculum preparation: Four fungal strains which have ability to produce lipase, were used throughout this study. Trichoderma viride SDTC EDF 002 and Aspergillus niger SDTC SRW-4 were obtained from Sime Darby Culture Collection Centre, Kuala Lumpur, Malaysia. Aspergillus terreus DSMZ 5770 and Aspergillus niger DSMZ 2466 were purchased from DSMZ Culture Collection Centre, Germany. The cultures were maintained in 20% (v/v) glycerol and preserved at -80°C. For spore production, the culture was grown in PDA Petri dish, incubated at 28±1°C for 7 days. The spores were harvested by pouring 10 mL of sterile 0.05% (v/v) Tween-20 into the Petri dish and dislodging the spores into suspension. The number of spore in suspension was determined using a Haemocytometer. For all strains, appropriate dilutions were made to obtain an inoculum with spore concentration of approximately 1x107 spores mL-1.
Solid state fermentation (SSF): Palm Kernel Cake (PKC), obtained from Premium Vegetables Company, Malaysia and was used as the main substrate in SSF for lipase production. These types of agro-wastes which once regarded as wastes are now known as the major renewable natural resource for various valuable products (Karmakar and Ray, 2010). The PKC was obtained by solvent extraction of the crushed palm kernel. This substrate was chosen due to its high nutrient composition (~13% made up of other chemicals like crude fat, crude protein, fibre, ether extract, ash and hemicelluloses). PKC (30 g) was placed in 500 mL flask and autoclaved at 121°C for 40 min. After cooling, 100% (v/w) of sterile distilled water and 10% (v/w) of inoculum were added into the flask. The content of the flask was mixed thoroughly and incubated at ambient temperature (~27±1°C) under static condition for 10 days. All fermentations, employing all four different fungal strains, were conducted concurrently in triplicates.
Extraction of lipase from solid substrate: At the specified incubation days (0, 2, 4, 6, 8 and 10 days), 15 mL of 0.02 M sodium phosphate buffer at pH 7.0 was added to 1.5 g of the fermented sample from each flask. The mixtures were shaken at 250 rpm and 4°C in a rotary shaker for 30 min. The mixtures were then filtered using Whatman filter paper (No. 1), where the filtrates were used for enzyme assays and determination of total protein content.
Analytical procedures: Total fungal biomass was estimated using glucosamine method as described by Swift (1973). In this method, 0.1 g of sample was added into 5 mL of 2 M HCl and heated at 95°C for 2 h. The aliquot (3 mL) was added with 1 mL of acetylacetone and heated at 95°C for 20 min. The solution was cooled and then added with 6 mL absolute ethanol and 1 mL of Ehrlich reagent. This mixture was incubated at 65°C for 15 min and the absorbance was read at 530 nm.
Lipase activity was determined using olive oil-polyvinyl alcohol emulsion as substrate (Mustranta et al., 1993). In this assay, 0.5 mL extracted sample was added into a reaction tube containing 2 mL sodium phosphate buffer (pH 7) and 2.5 mL olive-oil emulsion. The reaction was incubated at 37°C in water bath agitated at 200 rpm for 30 min. The reaction was terminated by the addition of 5 mL acetone-ethanol (1:1% v/v). The mixture was then titrated with 0.05 M NaOH using the autotitrator system (Mettler Toledo, T70) to obtain an equivalent point (EVP). One unit of lipase activity was defined as the amount of enzyme which liberated 1 μmol equivalent of fatty acid at 37°C, pH 7.0 in 1 min.
Protease activity was determined by casein digestion assay (Ibrahim-Granet and Bertrand, 1996). In this method, 0.5 mL 1% casein in 0.05 M Tris-phosphate buffer (pH 7.8) was equilibrated at 37°C for 5 min prior to the addition of 0.1 mL enzyme extract. The reaction mixture was incubated at 37°C, agitated at 200 rpm for 10 min. The reaction was stopped by adding 0.5 mL of 10% trichloroacetic acid and left to stand at room temperature for 10 min. The solution was centrifuged at 13,000 rpm for 20 min and the absorbance of the supernatant was then read at 280 nm using spectrophotometer (Thermo Electron Corporation, Biomate 5). One unit of protease activity was defined as the amount of enzyme which released acid soluble fragments equivalent to 0.001 A280 per minute at 37°C and pH 7.8.
Total protein content was determined using Bio-Rad Protein Assay Dye Reagent Concentrate (Chemoscience). In this method, 800 μL sample was added to 200 μL of the diluted Bradford reagent (Bradford reagent-H2O, 1:4% v/v) and then vortexed for 2 min. The solution was incubated at room temperature for 5 min and the absorbance was read at 595 nm.
Evaluation of growth kinetic: Growth and product formation kinetics were evaluated using logistic and Luedeking-Piret equations as described elsewhere (Gong and Lun, 1996; Younesi et al., 2005):
| (1) |
| (2) |
where, X = biomass concentration (mg g-1), t = time (h), μm = maximum specific growth rate (h-1), Xm = maximum biomass concentration (mg g-1), P = enzyme productivity (U g-1), α = growth associated product formation constant and β = non-growth associated product formation constant (1 h-1).
Microscopic imaging: The samples from SSF for the four fungal strains were withdrawn at different intervals (0, 2, 4, 6, 8 and 10 day) and examined under Scanning Electron Microscope (SEM) for imaging analysis. The fresh samples were viewed under Phillips SEM FEI Quanta 200 without any fixing or dehydration process. Micromorphology dimensions of spore diameter, hyphae diameter, branch diameter, tip diameter and tip length for each fungal sample were observed and recorded. However, hyphae length, branch length and number of tips per hyphae that has proven to increase protein production in submerged fermentation based on previous research work could not be observed from this study. This is because the fungal hyphae penetrates into the fermentation substrate and creates large complex mycelia surrounding the substrate that hinders the measurements to be taken.
Statistical analysis: Biomass content, enzyme productivity, total protein content and micromorphology data are means from triplicates. Micromorphology data for the zero and second day of fermentation could not be measured because distinct fungal structures were not detected in the samples. Generalizing the results from fungal fermentations is difficult due to their high sensitivity toward a slight variation in starting conditions, poor reproducibility and variations with strains (Cui et al., 1997). Therefore, statistical analysis was performed to obtain more comprehensive and useful information using the software package SPSS 15.0 for Windows (SPSS, Germany). The results presented were the mean values of ten replicates and the standard deviations were used to analyze the experimental data. Pearson correlation analysis was performed to measure the strength of association between two variables (bivariate data) statistically and also to evaluate whether the data are significant or not. Scatter plots are used to visualize the patterns of association between the bivariate data (independent and dependent variables). When the scatter plot shows a high level of association and correlation value, a regression equation was modeled in order to explain one variable to the other.
RESULTS
Relationship between fungal growth and enzyme formation rate: The performance and kinetic parameters values of solid state fermentation by several fungi for lipase production are shown in Table 1. A. niger SDTC SRW-4 had the highest lipase production (44.43 U g-1 DM) followed by A. niger DSMZ 2466 (42.05 U g-1 DM). The highest Xmax for all the four fungal strains was observed at day 6. Among the strains tested, the highest growth (Xmax = 0.710 mg g-1 DM), maximum specific growth rate (μmax = 0.029 h-1) and protease production (90.33 U g-1 DM) was obtained with A. terreus DSMZ 5770.
Figure 1 shows a typical time course of solid state fermentation by A. niger DSMZ 2466 for lipase production. Growth was in a lag phase up to day 2 of fermentation and exponential phase was continued until 6 days. The specific growth rate (μ) during exponential phase was 0.011 h-1 and growth entered a deceleration phase after 10 days. Lipase production increased gradually with fermentation time and the highest production rate (9.8693 U gh-1) was observed during exponential phase. Decrease in lipase production rate (2.4578 U gh-1) was observed when growth entered the deceleration phase (Table 2). The specific enzyme activity reached a maximum value (151.970 U mg-1 protein) after 10 days of fermentation. Luedeking-Piret equation (R2 = 0.8858) showed that the formation of lipase was partially growth associated, where the values of α and β were calculated as 39.665 and 1.0106, respectively.
Growth profile of T. viride STC EDF 002 was similar to A. niger DSMZ 2466. In contrary, lag phase was not observed for A. niger SDTC SRW-4 and A. terreus DSMZ 5770.
| Table 1: | Performance and kinetic parameters values of solid state fermentation using palm kernel cake for lipase production by several fungi |
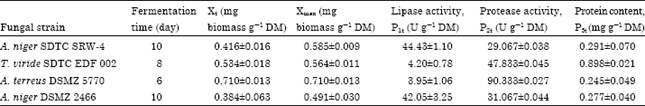 | |
| Data of each strain was selected from the highest lipase production from a ten-day fermentation time | |
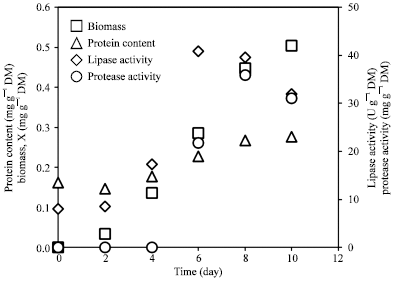 | |
| Fig. 1: | A typical time course of lipase production by A. niger DSMZ 2466 in solid state fermentation |
| Table 2: | Specific enzyme activity and product formation rate |
 | |
| * There were no distinct relationship between the product formation and growth of fungus based on the Luedeking-Piret equation | |
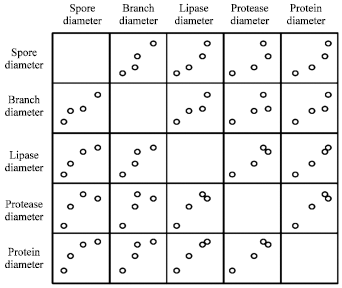 | |
| Fig. 2: | Scatter plots on high positive correlation bivariate data |
However, growth curve for all the fungal strains employed in this study showed that the exponential phase peaked at 6 days, showing that all the fungal strains had the same growth pattern on palm kernel cake and entered into a deceleration phase thereafter. During the 6 days of exponential growth, spore swelling, elongation of germ tubes and evolution of the total length of hyphae were observed. Subsequently, the water content in the fermentation substrate was reduced to low levels that caused substrate hardening. This hardening reduced and blocked the channeling of the substrate pores which in turn, retarded the penetration of the mycelia into the substrate to absorb the nutrients. The inability of the fungus to absorb more nutrients after hardening of substrate caused growth to enter into the deceleration phase. This was visually observed after 10 days of fermentation where approximately only 80% of the substrate was covered by fungal mycelia.
In fermentation with A. niger SDTC SRW-4, the highest specific activity of lipase (152.891 U mg-1 protein) was obtained after 10 days. In this case, lipase formation was found as growth-associated based on the Luedeking-Piret equation (R2 = 0.9773). Lipase formation and fungal growth for T. viride SDTC EDF 002 and A. terreus DSMZ 5770 did not show a similar trend and could not be correlated. Nevertheless, total protease production for T. viride SDTC EDF 002 contributed linearly to the total protein content in the fermentation substrate.
Micromorphology dimensions of fungal strains grown on PKC in SSF: Both A. niger strains of DSMZ 2466 and SRW-4 produced conidia black in colour, globose and with the diameter of 3.8 to 4.6 μm.
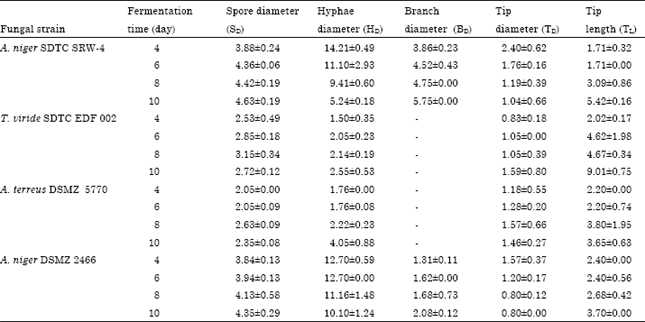
| Table 3: | Micromorphology dimensions of fungal strains grown on PKC in SSF |
 | |
On the other hand, A. terreus DSMZ 5770 produced smaller conidia (diameter of 2.0 to 2.6 μm) which were globose and have smooth surface while T. viride SDTC EDF 002 produced conidia (diameter of 2.5 to 3.2 μm) which was green in colour and grouped in sticky heads at the tips of the phialides.
Micromorphology dimensions for the spore diameter, hyphae diameter, branch diameter, tip diameter and tip length for the four fungal strains cultivated in solid state fermentation using PKC as substrate are summarized in Table 3. Data for the early stages of fermentation (0 to 2 days) could not be obtained because no distinct fungal spore structure could be identified. In addition, dimension of the branches diameter of T. viride SDTC EDF 002 and A. terreus DSMZ 5770 could not be measured because the fine branches overlapped and formed in a matrix structure.
A. niger DSMZ 2466 and A. niger SDTC SRW-4 displayed similar pattern on the morphology dimensions. In general, spore diameter and tip length were increased with fermentation time while reduction in hyphae diameter and tip length were observed. During growth in PKC, spore diameter and tip length for both fungal strains were more or less the same. However, hyphae and branch diameters for A. niger SDTC SRW-4 were comparatively larger than A. niger DSMZ 2466. Both fungal strains formed complex matrix structure within the substrate and produced long hyphae with diameter ranging from 9 to 12 μm.
Morphology dimension for T. viride SDTC EDF 002 and A. terreus DSMZ 5770 was almost similar, where the dimensions were increased with fermentation time. Spore diameter and tip length for T. viride SDTC EDF 002 were comparatively larger than those observed for A. terreus DSMZ 5770. On the other hand, hyphae and tip diameters for T. viride SDTC EDF 002 were smaller than the diameters for A. terreus DSMZ 5770.
Production of lipase by A. niger DSMZ 2466 and A. niger SDTC SRW-4 was about 10 fold higher than those obtained by A. terreus DSMZ 5770 and T. viride SDTC EDF 002. The ability of the fungi to produce lipase correlated well with the difference in their morphology dimensions.
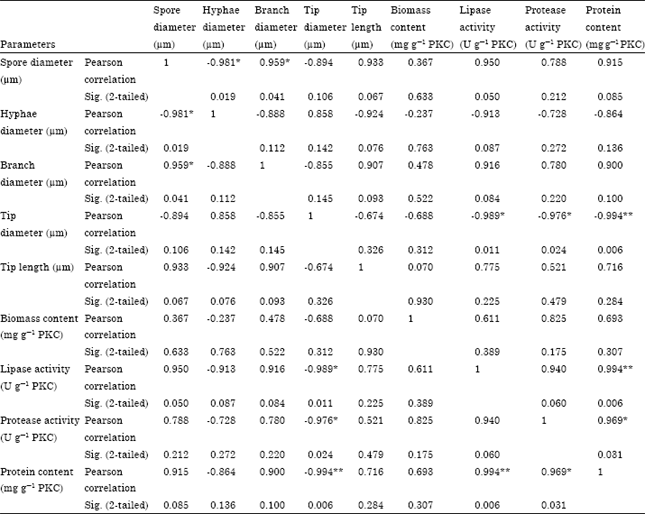
| Table 4: | Pearson correlation analysis for A. niger DSMZ 2466 |
 | |
| *Correlation is significant at the 0.05 level (2-tailed). **Correlation is significant at the 0.01 level (2-tailed) | |
Pearson correlation analysis for micromorphology dimensions and lipase production: Table 4 shows the variables, the significant levels and Pearson correlation for all dimensions of A. niger DSMZ 2466. Correlation measures the linear relationship between two quantitative variables which is known as bivariate data. Pearson correlation coefficient quantifies the strength of linear relationship between the bivariate data. The correlation coefficient ranges in value from -1 to 1. A value of 0 indicates no linear relationship between the variables while a value of +1 indicates a perfect positive relationship and -1 for a perfect negative relationship. This positive relationship among the bivariates can be visually demonstrated in a scatter plot to show the association between the two variables (Fig. 2). The following guidelines could be used to interpret correlation strength namely small, medium and large with the values of 0.10 to 0.29, 0.30 to 0.49 and 0.50 to 1.0, respectively (Cohen, 1988).
The data which have high significant levels (p<0.05) are extracted for further analysis to ensure the data are normally distributed (Table 4). Pearson correlation for lipase activity versus protein content (R2 = 0.994) and spore diameter versus branch diameter (R2 = 0.959) have high significance levels, suggesting the occurrence of a large strength correlation between these variables. Bivariate data of spore diameter and lipase activity was an exceptional data with the significance level on par at 0.05 with a high correlation coefficient (0.950).
Spore diameter versus hyphae diameter and tip diameter versus lipase activity, protease activity and protein content showed very strong negative relationship at a high significance level (R2>0.97). These results indicate that these variables are independent. For example, size of the spore was not influenced by the thickness of the hyphae. The thickness of the tip also did not influence lipase and protease production.
Both, lipase and protease activity for A. niger SDTC SRW-4 was strongly correlated with the size of the spore and the thickness of the branch. Moreover, production of these enzymes had similar trends throughout the fermentation period. Pearson correlation showed that the thickness of the hyphae and tip did not influence lipase production. Strong negative relationship was observed between variables of hyphae and branch diameters; and spore and tip diameters. Spore diameter, branch diameter and lipase production in fermentation with A. niger SDTC SRW-4 and A. niger DSMZ 2466 were interrelated as evaluated by Pearson correlation. The bivariate significance level for branch diameter and lipase activity for A. niger DSMZ 2466 was low (>0.05). However, the correlation was high (R2>0.9) showing a positive relationship.
T. viride SDTC EDF 002 and A. terreus DSMZ 5770 showed positive correlations for tip length and tip diameter as well as size of hyphae diameter and total protein content in the fermentation substrate. Linear relationship with the tip length and tip diameter was observed for spore size of A. terreus DSMZ 5770. Even though lipase and protease production by A. terreus DSMZ 5770 shared similar pattern with A. niger SRW-4 but Pearson correlation did not show any inter-relationship between the micro-morphology dimensions with lipase production.
Regression analysis: Regression analysis creates a model that relates a dependent (outcome or response) variable to an independent variable. The appropriate model is subsequently used for prediction of equations. Table 5 describes the summary of the models that were generated from a curve fit plot. Linear or quadratic models were selected based on their best regression value that fits the curve plot. Figure 3 demonstrates an example of a curve fit plot of A. niger DSMZ 2466 spore diameter (independent variable) versus lipase activity (dependent variable), showing that the quadratic curve best-fitted to the plot with R2 of 0.999.
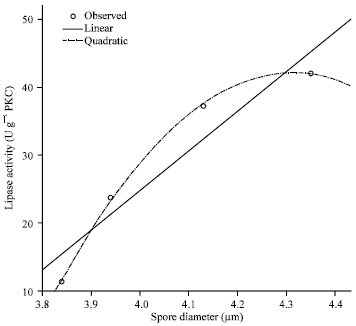 | |
| Fig. 3: | Example of a curve fit plot from a regression analysis (data extracted from A. niger DSMZ 2466) |
| Table 5: | Summary of regression analysis and equation models |
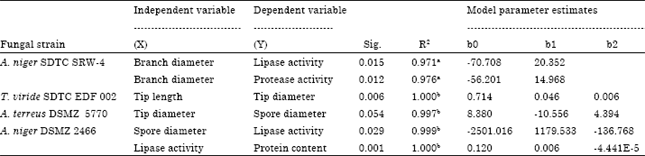 | |
| Only bivariate data which are significant (<0.05) and R2 more than 0.97 are tabled. aLinear model equation is Y = b0 + (b1 * X). The series values are modeled as a linear function of time. bQuadratic model equation is Y = b0 + (b1 * X) + (b2 * X2) | |
Both strains of A. niger DSMZ 2466 and A. niger SDTC SRW-4 tested in this study showed good micro-morphology characteristics with larger spore sizes and hyphae diameter as compared to A. terreus DSMZ 5770. Lipase activity and the branch diameter for A. niger SDTC SRW-4 was highly correlated in a linear model (R2 = 0.971) while lipase activity and spore diameter for A. niger DSMZ 2466 was correlated in a quadratic model (R2 = 0.999). For A. niger DSMZ 2466, the total protein content was strongly influenced by the total lipase activity in the fermentation substrate. There was no relationship between the micro-morphology dimensions and production of lipase for T. viride SDTC EDF 002 and A. terreus DSMZ 5770. Nevertheless, the relationships between tip length and tip diameter as well as the tip diameter and spore diameter were existence.
DISCUSSION
Palm oil kernels are available as palm kernel press cake (PKC) which is a residue from extraction of oil from palm kernels. The extraction can be either mechanical or solvent extraction resulting in a residue containing about 50% carbohydrate and 15-20% protein (Knudsen, 1997; Cervero et al., 2010). Since PKC consisted of sugar such as mannose (30-35%) and glucose (7-9%) in PKC (Dusterhoft et al., 1991; Knudsen, 1997), it has potential to be used as substrate in SSF for production of industrial enzymes. All the four fungal strains tested in this study showed good growth in SSF using PKC as substrate, where final biomass content ranging from 0.491 to 0.710 mg biomass g-1 DM was achieved. All the strains grew in filamentous form and the hyphae showed ability to elongate and form complex branches filling spaces in the substrate matrix at a similar rate in the exponential phase. This fungus utilizes the nutrients available through the development of penetrative hyphae into the substrate matrix and degrades the high content of hemicelluloses and celluloses by producing various types of hydrolytic enzymes. The predominance between the ability to penetrate and to degrade may be related to the extent of diffusion and assimilation of nutrients from the medium to fungus which is associated to different control mechanisms related to growth kinetics (Cano and Bago, 2005; Boswell, 2008).
A. niger SDTC SRW-4 and A. niger DSMZ 2466 showed high productivity of lipase enzyme while T. viride SDTC EDF 002 and A. terreus DSMZ 5770 showed higher productivity of protease enzyme. Several other types of hydrolytic enzymes such as xylanases, mannanases or cellulases were also produced by these fungal strains during growth on PKC due to high content of mannan (~35.2%) (Cervero et al., 2010).
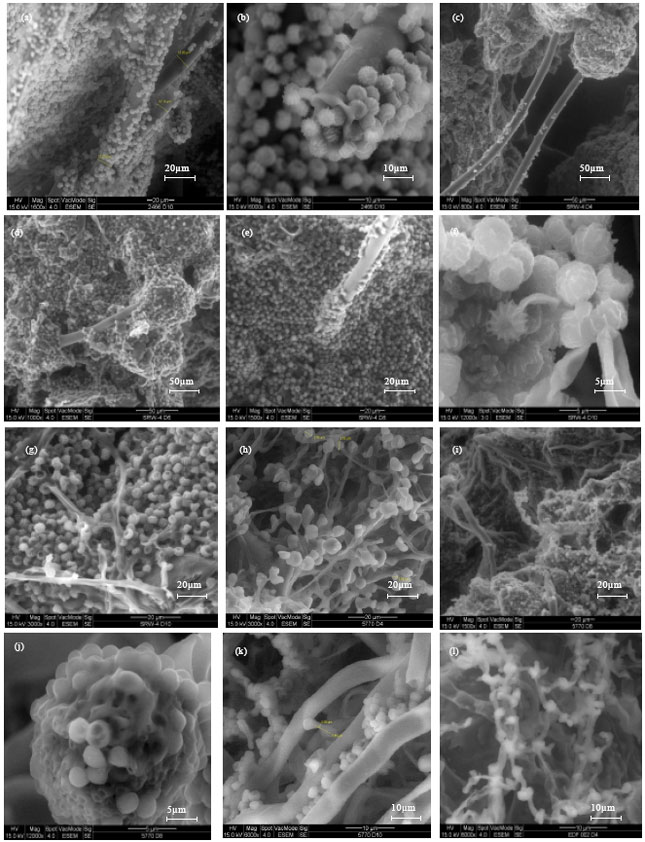  | |
| Fig. 4(a-o): | SEM images on growth of fungal strains on PKC, (a-b) A. Niger DSMZ 2466, (c-g) A. Niger SRW-4, (h-k) A. terreus DSMZ 5770 and (l-o) T. viride EDF 022 |
Two stages of growth were occurred in SSF inoculated with spores; (1) swelling of a population of conidiospores and (2) full development of branched hyphae. Result from this study indicated that increased in mean equivalent spore diameter was approximately linear with time. Barry et al. (2009) has stressed that differences in spore swelling rate and germination time would make a significant contribution to the observed variation in the sizes. Both fungal strains, A. niger SDTC SRW-4 and A. niger DSMZ 2466, have higher spore swelling and germination rate as compared to T. viride SDTC EDF 002 and A. terreus DSMZ 5770 which have higher ability in producing lipase. The presence of a layer of mannose-rich glycoproteins in the walls of A. niger germinating spores was associated with adhesion phenomena (Brul et al., 1997). Large diameters of germinating spores contained large amount of mannose-rich glycoproteins layer existed which may related to higher enzyme production.
From SEM results in Fig. 4, a complex branching with fine branches emerged from the hyphae was observed for T. viride SDTC EDF 002 and A. terreus DSMZ 5770. The formation of this matrix made the dimensions of branch length and diameter difficult to be measured. Although, the spore diameter and hyphae diameter of these two fungal strains were shorter as compared to A. niger SDTC SRW-4 and A. niger DSMZ 2466, they have good growth on PKC. The local fractal dimension within a colony of T. viride was increased with branching frequency which can be seen from the occurrence of ‘loops’ in the mycelium (Hitchcock et al., 1996). In addition, good growth is imputable from the branching mechanism that promotes exponential growth of filamentous microorganisms and together with tip extension which determines the overall specific growth rate and the morphology of freely dispersed hyphal elements (Pazouki and Panda, 2000). The branching mechanism in T. viride SDTC EDF 002 and A. terreus DSMZ 5770 has no relationship with high lipase production but may related to protease production. Results from this study showed that large spore and branch diameter was correlated with high ability in producing lipase, as observed in models for A. niger SDTC SRW-4 and A. niger DSMZ 2466. A good linear relationship between sporangium diameter and enzyme production using Pearson correlation analysis has also been reported (De Nicolas-Santiago et al., 2006).
Hyphae and tip diameters for A. niger SDTC SRW-4 and A. niger DSMZ 2466 strains were decreased with fermentation time, most probably due to differentiation. In the production of secondary metabolites by Streptomyces differentiation was observed for growth on solid media (Pons et al., 1998). Vegetative mycelia consist of thick filaments and with aging, septation occurs and the filaments get thinner or tend to lose their cellular content while giving rise to empty compartments. The empty parts then start to appear at the end of the rapid growth phase and continue throughout the fermentation where initially the empty parts appeared to the tips of the filaments. This could also be related to changes in the membrane permeability which induces a leakage of intracellular materials. The high degree of vacuolation may also be viewed as a normal development change in the culture under imperfect conditions for fungal growth, for example O2 limitation (Paul et al., 1994).
CONCLUSION
All lipase-producing fungal strains (A. niger DSMZ 2466, A. niger SDTC SRW-4, T. viride SDTC EDF 002 and A. terreus DSMZ 5770) employed in this study showed variation in the micro-morphology characteristics towards enzyme production though the growth patterns on PKC were more or less the same. In general, hyphae and spore diameters for A. niger were larger as compared to A. terreus and T. viride which was about 7-fold and 2-fold higher, respectively. Strong positive relationship between spore and branch diameters with lipase production was observed for fermentations with A. niger. On the other hand, correlation between morphology dimensions with lipase activity was not observed for fermentations with A. terreus and T. viride. However, the tip length and tip diameter had a strong influential relationship for T. viride while the tip diameter and spore diameter showed strong positive relationship for A. terreus.
ACKNOWLEDGMENTS
This research work was financially supported by Sime Darby Technology Centre Sdn. Bhd. Ms. Anusha Nair would also like to extent appreciation to Sime Darby Technology Centre Sdn. Bhd. for her PhD scholarship.
REFERENCES
- Ahamed, A. and P. Vermette, 2009. Effect of culture medium composition on Trichoderma reesei's morphology and cellulase production. Bioresour. Technol., 100: 5979-5987.
CrossRef - Barry, D.J., C. Chan and G.A. Williams, 2009. Morphological quantification of filamentous fungal development using membrane immobilization and automatic image analysis. J. Ind. Microbiol. Biotechol., 36: 787-800.
CrossRefDirect Link - Boswell, G.P., 2008. Modelling mycelial networks in structured environments. Mycol. Res., 112: 1015-1025.
CrossRef - Brul, S., A. King, J.M. van der Vaart, J. Chapman, F. Klis and C.T. Verrips, 1997. The incorporation of mannoproteins in the cell wall of S. cerevisiae and filamentous Ascomycetes. Antonie Van Leeuwenhoek, 72: 229-237.
CrossRef - Cano, C. and A. Bago, 2005. Competition and substrate colonization strategies of three polyxenically grown arbuscular mycorrhizal fungi. Mycologia, 97: 1201-1214.
CrossRef - Cervero, J.M., P.A. Skovgaard, C. Felby, H.R. Sorensen and H. Jorgensen, 2010. Enzymatic hydrolysis and fermentation of palm kernel press cake for production of bioethanol. Enzyme Microb. Technol., 46: 177-184.
CrossRef - Cui, Y.Q., G.G.J.M. van der Lans and K.C.A.M. Luyben, 1997. Effect of agitation intensities on fungal morphology of submerged fermentation. Biotechnol. Bioeng., 55: 715-726.
CrossRef - De Nicolas-Santiago, S., C. Regalado-Gonzalez, B. Garcia-Almendarez, F.J. Fernandez, A. Tellez-Jurado and S. Huerta-Ochoa, 2006. Physiological, morphological and mannanase production studies on Aspergillus niger uam-gs1 mutants. Elect. J. Biotechnol., 9: 51-60.
CrossRefDirect Link - Dusterhoft, E.M., A.G.J. Voragen and F.M. Engels, 1991. Non-starch polysaccharides from sunflower (Helianthus annuus) meal and palm kernel (Elaeis guineenis) meal-preparation of cell wall material and extraction of polysaccharide fractions. J. Sci. Food Agric., 55: 411-422.
CrossRefDirect Link - Freire, D.M., E.M.F. Teles, E.P.S. Bon and G.L. Sant' Anna Jr., 1997. Lipase production by Penicillium restrictum in a bench-scale fermenter: Effect of carbon and nitrogen nutrition, agitation and aeration. Applied Biochem. Biotechnol., 63: 409-421.
CrossRefPubMedDirect Link - Gong, H. and S. Lun, 1996. The kinetics of lysine batch fermentation. Chin. J. Biotechol., 12: 219-225.
Direct Link - Gordon, C.L., D.B. Archer, D.J. Jeenes, J.H. Doonan, B. Wells, A.P.J. Trinci and G.D. Robson, 2000. A glucoamylase: GFP gene fusion to study protein secretion by individual hyphae of Aspergillus niger. J. Microbiol. Methods, 42: 39-48.
CrossRef - Grimm, L.H., S. Kelly, R. Krull and D.C. Hempel, 2005. Morphology and productivity of filamentous fungi. Applied Microbiol. Biotechnol., 69: 375-384.
CrossRef - Haack, M.B., L. Olsson, K. Hansen and A.E. Lantz, 2006. Change in hyphal morphology of Aspergillus oryzae during fed-batch cultivation. J. Applied Microbiol. Biotechnol., 70: 482-487.
CrossRef - Hitchcock, D., C.A. Glasbey and K. Ritz, 1996. Image analysis of space-filling by networks: Application to a fungal mycelium. Biotechnol. Tech., 10: 205-210.
CrossRef - Hwang, H.J., S.W. Kim, C.P. Xu, J.W. Choi and J.W. Yun, 2004. Morphological and rheological properties of the three different species of basidiomycetes Phellinus in submerged cultures. J. Applied Microbiol., 96: 1296-1305.
CrossRef - Ibrahim-Granet, O. and O. Bertrand, 1996. Separation of proteases: Old and new approaches. J. Chromatogr. B: Biomed. Sci. Appl., 684: 239-263.
CrossRef - Karmakar, M. and R.R. Ray, 2010. Extra cellular endoglucanase production by rhizopus oryzae in solid and liquid state fermentation of agro wastes. Asian J. Biotechnol., 2: 27-36.
CrossRef - Knudsen, K.E.B., 1997. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Technol., 67: 319-338.
CrossRefDirect Link - Lim, J.S., J.H. Lee, J.M. Kim, S.W. Park and S.W. Kim, 2006. Effects of morphology and rheology on neo-fructosyltransferase production by Penicillium citrinum. Biotechnol. Bioproc. Eng, 11: 100-104.
Direct Link - Mahadik, N.D., U.S. Puntambekar, K.B. Bastawde, J.M. Khire and D.V. Gokhale, 2002. Production of acidic lipase by Aspergillus niger in solid state fermentation. Process Biochem., 38: 715-721.
CrossRefDirect Link - Muller, C., M. McIntyre, K. Hansen and J. Nielsen, 2002. Metabolic engineering of the morphology of Aspergillus oryzae by altering chitin synthesis. J. Applied Environ. Microbiol., 68: 1827-1836.
CrossRefPubMedDirect Link - Mustranta, A., P. Forssell and K. Poutanen, 1993. Applications of immobilized lipases to transesterification and esterification reactions in nonaqueous systems. Enzyme Microb. Technol., 15: 133-139.
Direct Link - Oncu, S., C. Tari and S. Unluturk, 2007. Effect of various process parameters on morphology, rheology and polygalacturonase production by Aspergillus sojae in a batch bioreactor. Biotechnol. Prog., 23: 836-845.
CrossRef - Paul, G.C., C.A. Kent and C.R. Thomas, 1994. Image analysis for characterizing differentiation of Penicillum chrysogenum. Food Bioprod. Process., 72: 95-105.
Direct Link - Pazouki, M. and T. Panda, 2000. Understanding the morphology of fungi. Bioprocess Biosyst. Eng., 22: 127-143.
CrossRef - Pons, M.N., J.F. Drouin, L. Louvel, B. Vanhoutte, H. Vivier and P. Germain, 1998. Physiological investigations by image analysis. J. Biotechnol., 65: 3-14.
CrossRef - Spohr, A., M. Carlsen, J. Nielsen and J. Villadsen, 1997. Morphological characterization of recombinant strains of Aspergillus oryzae producing α-amylase during batch cultivations. Biotechnol. Lett., 19: 257-262.
CrossRef - Swift, M.J., 1973. The estimation of mycelial biomass by determination of the hexosamine content of wood tissue decayed by fungi. Soil Biol. Biochem., 5: 321-332.
CrossRefDirect Link - Teng, Y., Y. Xu and D. Wang, 2009. Changes in morphology of Rhizopus chinensis in submerged fermentation and their effect on production of mycelium-bound lipase. J. Bioprocess Biosyst. Eng., 32: 397-405.
CrossRef - Wessels, J.G.H., 1993. Wall growth protein excretion morphogenesis in fungi. New Phytol., 123: 397-413.
CrossRef - Wosten, H.A.B., S.M. Moukha, J.H. Sietsma and J.G.H. Wessels, 1991. Localization of growth and secretion of proteins in Aspergillus niger. J. Gen. Microbiol., 137: 2017-2023.
CrossRefPubMedDirect Link - Younesi, H., G. Najafpour and A.R. Mohamed, 2005. Ethanol and acetate production from synthesis gas via fermentation processes using anaerobic bacterium, Clostridium ljungdahlii. Biochem. Eng., 27: 110-119.
CrossRef