
Mukesh
Wildlife Institute of India, P.O. Box 18, Chandrabani, Dehradun 248 001, Uttarakhand, India
Rajiv S. Kalsi
M.L.N. College, Yamuna Nagar 135 001, Haryana, India
R.P. Mandhan
Department of Biotechnology, Kurukshetra University, Kurukshetra-136119, Haryana, India
S. Sathyakumar
Wildlife Institute of India, P.O. Box 18, Chandrabani, Dehradun 248 001, Uttarakhand, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 3 | Page No.: 293-301







ABSTRACT
Genetic diversity of Red Junglefowl (Gallus gallus) in five range states of northern India was explored to make conservation action plan for genetically endangered bird species. A total of 111 Red Junglefowl samples were collected and genotyped with 15 polymorphic microsatellite loci. Altogether 173 alleles were found with mean (±SE) allelic number of 11.53±6.96. Loci LEI0234 yielded the highest number of alleles (Na = 32) while loci LEI0166, MCW0222 and MCW0098 yielded the lowest number of alleles (Na = 5) in the population. Out of 15 microsatellite loci used, 14 loci showed PIC value higher than 0.5 and considered informative for population genetic studies. The observed heterozygosity (Ho) in the population ranged between 0.378 and 0.637, with mean (±SE) of 0.391±0.134. Hardy-Weinberg equilibrium (HWE) estimations were conducted and all the 15 loci were found to follow the equation (p>0.05). No evidence for linkage disequilibrium was observed among pair of loci. Genetic bottleneck hypotheses were also tested, suggesting that RJF has not experienced a genetic bottleneck in the recent past. Persent study revealed the considerable genetic variation in this population, which is an important factor for genetic resources conservation, evaluation and for conservation breeding.
PDF Abstract XML References Citation
Received: September 20, 2010;
Accepted: December 21, 2010;
Published: February 07, 2011
How to cite this article
Mukesh, Rajiv S. Kalsi, R.P. Mandhan and S. Sathyakumar, 2011. Genetic Diversity Studies of Red Junglefowl Across its Distribution Range in Northern India. Asian Journal of Biotechnology, 3: 293-301.
DOI: 10.3923/ajbkr.2011.293.301
URL: https://scialert.net/abstract/?doi=ajbkr.2011.293.301
DOI: 10.3923/ajbkr.2011.293.301
URL: https://scialert.net/abstract/?doi=ajbkr.2011.293.301
INTRODUCTION
The wild Red Junglefowl (Gallus gallus) (RJF) is the wild ancestor of all domestic chicken in the world and is one of the most important species to mankind due to the economic and cultural significance to human civilization (Fumihito et al., 1996). The present day multi-billion dollar poultry industry is based on the wild RJF and will depend on it in future as well. India is the fifth largest egg producer with over 40 billion eggs and over 650 million broilers (www.fao.org). The evolution of chicken can be conceived as a three steps processes-evolution and speciation of wild ancestor; domestication of wild species, and diversification into numerous varieties under artificial selection by humans. Exotic high yielding breeds have replaced our native breeds. The populations of domestic animals and their wild ancestors provide a valuable source of genetic diversity that may be exploited to develop animal models for quantitative traits of biological and medicinal interest. Conservation of genetically pure wild forms or their representatives have great potential to make significant contribution to the study of some economically important genetic traits of domestic forms. Unlike other domestic species, where the ancestor from which the present day animals evolved do not exist, the domestic chicken offers a unique system to compare the effect of natural selection and therefore it has immense value. Hanotte and Jianlin (2005) stated that the reservoir of genetic diversity the putative wild ancestors’ are now either extinct or low in numbers and threatened by extinction. Although RJF is considered abundant both in captivity and in wild but due to hybridization with feral or domestic chicken, the wild RJF is believed to be threatened with genetic endangerment throughout its distribution range (Peterson and Brisbin, 1998). Habitat loss, fragmentation, degradation and poaching have also threatened the wild RJF populations in India. Recent studies (Brisbin and Peterson, 2007; Kanginakudru et al., 2008) suggest that phenotypic characters may have little, if any, utility in characterising RJF stock as to their genetic purity. To approach these questions of genetic purity, however apart from phenotypic markers, additional tools drawn from molecular genetics have yet to be applied. Currently, microsatellites are widely used since they are numerous, randomly distributed in the genome, highly polymorphic and show co-dominant inheritance (Karaca et al., 1999). Many microsatellites have recently become available in chicken and have been mapped in reference populations (Crooijmans et al., 1996a; Groenen et al., 2000). These markers provide a powerful tool for quantitative trait loci (QTL) research and have also been successfully used to study the genetic relationship between and within chicken populations (Vanhala et al., 1998; Ponsuksill et al., 1999; Zhou and Lamont, 1999; Romanov and Weigend, 2001 Rosenberg et al., 2001).
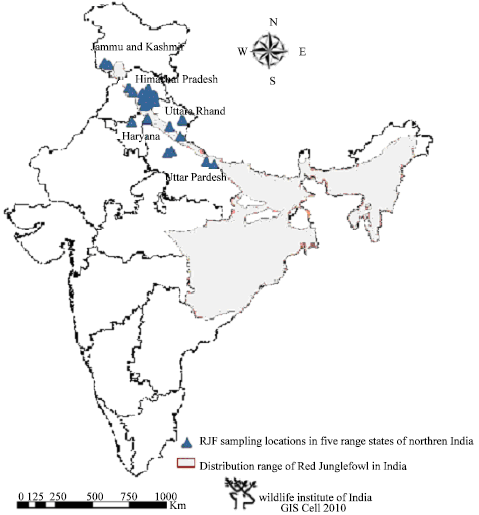 | |
| Fig. 1: | Map showing Red Junglefowl sampling location in northern India along with its distribution in five different zones in India |
Reliable information on allele frequencies was obtained from chicken blood or DNA pools using minisatellite markers (Dunnington et al., 1994) as well as microsatellites (Khatib et al., 1994, Crooijmans et al., 1996b, Tadano et al., 2007, Bao et al., 2007, Mukesh et al., 2010a). Some research on origin, phylogeny, genetic diversity and domestication of RJF (Fumihito et al., 1996; Vanhala et al., 1998; Romanov and Weigend, 2001; Niu et al., 2002; Moiseyeva et al., 2003; Hillel et al., 2003; Olowofeso et al., 2005; Ya-Bo et al., 2006; Sharma, 2006; Kanginakudru et al., 2008; Hassen et al., 2009) have been carried out. The RJF is widely distributed and its five subspecies are spread from the Indian subcontinent eastwards across Myanmar, South China, Indonesia to Java (Johnsgard, 1986). In India, two sub-species occur, the type specimen, Gallus gallus murghii and Gallus gallus spadiceus (Fernandes et al., 2009). While the former is found in the north and central part of India, extending eastwards to Orissa and West Bengal, the latter is confined to the northeastern parts of India. In order to obtain information on the current status and distribution of RJF in India, extensive surveys were carried out in different range states that fall under five regions viz., Northern, Central, Eastern, Southeastern and Northeastern (Fig. 1). The species is present in 205 Districts in 21 range states in India. Of the 255 Protected Areas that occur within the RJF distribution range in India, 190 (31 National Parks and 159 Wildlife Sanctuaries) have reported presence in their areas (Fernandes et al., 2009).
MATERIALS AND METHODS
Live-trapping, sample collection and DNA extraction: In total, 111 RJF samples (16 from wild and 95 from captivity) were collected from five range states of northern India (Table 1). RJF were live-trapped across its distribution range states in northern India, which includes Jammu and Kashmir, Himachal Pradesh, Haryana, Uttarakhand and Uttar Pradesh. We also collected blood/feather samples from captive RJF stock in zoos/pheasantries in these States. In addition, the efficacy of different sampling methods and analytical techniques for g DNA extraction with considering cost and efforts have also been conducted (Mukesh et al., 2010b). Approximately 500 μL blood was drawn from live-trapped bird and stored in DNA zol BD as well as on FTA cards (Mackey et al., 1996, 1998).
| Table 1: | Details of Red Junglefowl samples collected from 5 distribution range states in northern India |
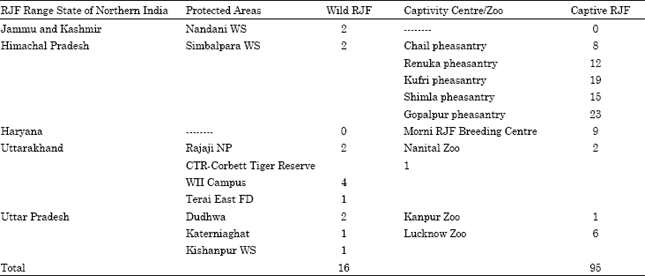 | |
Freshly pulled primary feathers were preserved in 70% alcohol (www.gallus.forestry.uga.edu/ggg/protocols/pdf/feather.pdf) while moulted feathers and hatched egg shell were stored in an envelopes/zip lock bag (www.absc.usgs.gov/). Genomic DNA was extracted from blood following DNA zol BD based protocol while DNA was extracted from tissue, feathers and egg shell membrane using Qiagen DNeasy tissue kit, following the manufacturer’s instructions. This present study is the part of a national project titled ‘Conservation of Red Junglefowl in India that is being carried out at the Wildlife Institute of India to develop an action plan for conserving the wild ancestor in its natural habitat. The project was initiated in September 2006 and will conclude by February 2010. Samples are being collected from its potential distribution range in 21 states of India.
Microsatellite genotyping: A set of 15 polymorphic microsatellite markers, developed for chicken and earlier used in the (AVIANDIV et al., 2000) project were used in the study (Table 2) (www.aviandiv.tzv.fal.de/primer_table.html). The PCRs were carried out in 10 μL reaction volume in an Applied biosystem thermal cycler (2700 and 2720), containing 1 X PCR Buffer (50 mM KCl, 10 mM Tris-HCl), 1.5 mM MgCl2, 200 μM of each d-NTP, 1.25 μg BSA, 4 p-mole of each primer, 0.5 unit of Taq DNA polymerase, 50 to 80 ng of gDNA. The amplification conditions were: 2 min initial denaturation at 94°C, followed by 35 cycles of denaturation at 94°C for 45 sec, annealing at specific temperature (Table 2) for 45 sec, and extension at 72°C for 2 min with a final extension at 72°C for 15 min. 5 μL of PCR products were mixed with 1 μL of loading buffer and then loaded onto a 2% agarose gel containing ethidium bromide and visualized over UV light to detect amplification. PCR products were pooled and denatured at 95°C for 5 min and microsatellite genotyping was carried out using ABI 3130 Genetic Analyser (Applied Biosystem) with GeneScan-500 (-250) LIZ as the internal lane size standard. Data were collected and analysed using GeneMapper Software (Version 3.7, Applied Biosystem) (Estes-Zumpf et al., 2010).
| Table 2: | Description of Microsatellite Loci used in the study |
 | |
Genetic diversity analysis: Genetic diversity estimates i.e., Observed and expected heterozygosity, observed and effective number of alleles were carried out after Nei (1972, 1973) using POPGENE software Francis et al., 1999). Using allelic frequencies, polymorphic information content (PIC), a measure of marker’s informativeness, was calculated with the Cervus (ver. 3.0) computer program (Kalinowski et al., 2007). Deviation from Hardy-Weinberg equilibrium was tested using the exact test of POPGENE (Francis et al., 1999). Heterogeneity of deviations from Hardy-Weinberg equilibrium among the microsatellite loci was investigated by considering the deviations as correlation coefficients and tested accordingly (Barker et al., 2001). Heterozygote deficiencies were estimated as FIS = (Ho- He)/He, where Ho and He are the observed and expected frequency of heterozygotes, respectively. Linkage disequilibrium (LD) test between pairs of loci was performed in FSTAT version 2.9.3 (Goudet, 1995). Evidence of a recent genetic bottleneck was tested using the program BOTTLENECK (Piry et al., 1999).
RESULTS AND DISCUSSION
Polymorphism of markers and genetic diversity statistics: In this study, RJF population of northern India showed enough genetic polymorphism. All 111 RJF samples were successfully amplified and 173 distinct alleles were distinguished over the 15-microsatellite loci used. Linkage disequilibrium was not detected between the investigated loci. Therefore all the loci were retained for the further analysis. Genetic diversity measures of RJF population are presented in Table 3. The number of observed alleles were ranged from 5 for loci LEI0166, MCW0222, MCW0098 and 32 for locus LEI0234 while the overall mean number of alleles per locus was reported 11.53 (±6.9 SE). The effective number of alleles were ranged from 2.31 for locus MCW0098 to 10.05 for locus LEI0234 with mean (±SE) of 4.67±2.10 (Table 3). The observed number of alleles for all the 15 loci exceeded the effective number of alleles. All the microsatellite loci showed PIC values higher than 0.5 except MCW0098 (PIC-0.472) which is normally considered as informative in population-genetic analyses (Botstein et al., 1980). The mean PIC in the present study was 0.7119. Mean (±SE) observed heterozygosity over the 15 loci was 0.391±0.134, which was lower than the expected heterozygosity. The expected heterozygosity within RJF population of northern India ranged from 0.570 (MCW0098) to 0.905 (LEI0234) with overall mean (±SE) of 0.755±0.090. Thus RJF population of Northern India gave the impression of high-quality reservoir of genetic variation which also corroborates with our own study carried out in state of Himachal Pradesh with captive samples (Mukesh et al., 2010a). Our within-population inbreeding estimate (FIS) was significantly positive, as derived from table-wide randomizations (p<0.05). The FIS estimates ranged between 0.230 for locus MCW0081 to 0.755 for locus MCW0037. The mean FIS estimate was 0.478. Heterozygosity, however, is not reduced proportionally, because rare alleles contribute little to heterozygosity. Recent genetic bottleneck was tested using the statistical test based on the difference between allelic diversity and heterozygosity (Piry et al., 1999).The allele frequency spectrum was visualized by the qualitative graphical method as described by Cornuet and Luikart (1996) (Fig. 2). The microsatellite alleles were arranged into 10 frequency classes, which make sure whether the distribution followed the normal L-shaped form, where alleles with low frequencies (0.01-0.1) are the most abundant. The observed allelic distribution revealed that the population did not encounter a genetic bottleneck in the recent past. The significant level of genetic variability in RJF population of northern India, notwithstanding its diminished population size, is indicative of a valuable reservoir of genetic diversity in this population.
| Table 3: | Genetic Diversity statistics of Red Junglefowl in northern India |
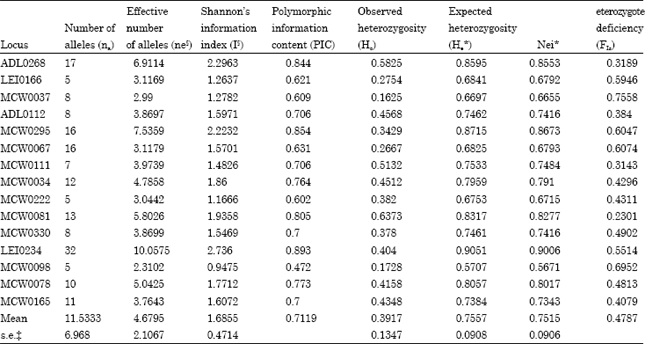 | |
| §ne: effective number of alleles (Kimura and Crow 1964), I: Shannon’s information index (Lewontin 1972); * :He :Expected heterozygosity was cmputed using Levene (1949) and Nei (1973) epected heterozygosity, ‡ s.e.:- Standard error | |
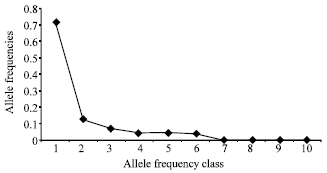 | |
| Fig. 2: | Normal ‘L-shaped mode-shift’ graph showing lack of recent genetic bottleneck in Red Junglefowl population in northern India |
CONCLUSION
The high genetic diversity in this population is important for genetic resources conservation and utilization. The information from the DNA markers together with phenotypic performance and population history provides reliable guidelines that can be used in developing practical strategies for conservation purposes and for breeding programs. High-priority action plan has to be initiated and the villagers especially tribal should be educated for comprehensive safeguarding of wild RJF by not selectively removing males from wild populations for upgrading their domestic breeds which may further weaken the diversity levels through limitation of wild mating males.
ACKNOWLEDGMENTS
The study was funded by Wildlife Institute of India, Dehradun, India. We acknowledge Director, Dean and Dr. S.P. Goyal, WII for their consistent encouragement and support. We thank Chief Wildlife Wardens and Forest Officers in RJF range states of northern India. At WII, our special thanks are due to Sudhanshu Mishra, Lalit K. Sharma and Manish Bhardwaj for their valuable discussions. We gratefully acknowledge the help received from Merwyn Fernandes during sample collection and the lab colleagues during wet lab operations.
REFERENCES
- Botstein, D., R.L. White, M. Skolnick and R.W. Davis, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet., 32: 314-331.
PubMedDirect Link - Brisbin Jr., L. and A.T. Peterson, 2007. Playing chicken with red junglefowl: Identifying phenotypic markers of genetic purity in Gallus gallus. Anim. Conserv., 10: 429-435.
CrossRefDirect Link - Cornuet, J.M. and G.L. Luikart, 1996. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics, 144: 2001-2014.
PubMed - Crooijmans, R.P., A.F. Groen, A.J. van Kampen, S. van der Beek, J.J. van der Poel and M.A. Groenen, 1996. Microsatellite polymorphism in commercial broiler and layer lines estimated using pooled blood samples. Poult. Sci., 75: 904-909.
PubMed - Crooijmans, R.P., P.A. van Oers, J.A. Strijk, J.J. van der Poel and M.A. Groenen, 1996. Preliminary linkage map of the chicken (Gallus domesticus) genome based on microsatellite markers: 77 new markers mapped. Poult. Sci., 75: 746-754.
PubMedDirect Link - Dunnington, E.A., L.C. Stallard, J. Hillel and P.B. Siegel, 1994. Genetic diversity among commercial chicken populations estimated from DNA fingerprints. Poult. Sci., 73: 1218-1225.
PubMedDirect Link - Estes-Zumpf, W.A., J.L. Rachlow, L.P. Waits and K.I. Warheit, 2010. Dispersal, gene flow and population genetic structure in the pygmy rabbit (Brachylagus idahoensis). J. Mammol., 91: 208-219.
Direct Link - Fernandes, M., Mukesh, S. Sathyakumar, R. Kaul, R.S. Kalsi and D. Sharma, 2009. Conservation of red junglefowl Gallus gallus in India. Int. J. Galliformes Conserv., 1: 94-101.
Direct Link - Fumihito, A., T. Miyake, M. Takada, R. Shingu and T. Endo et al., 1996. Monophyletic origin and unique dispersal patterns of domestic fowls. Proc. Natl. Acad. Sci. USA., 93: 6792-6795.
PubMedDirect Link - Goudet, J., 1995. FSTAT (version 1.2): A computer program to calculate F-statistics. J. Heredity, 86: 485-486.
Direct Link - Groenen, M.A., H.H. Cheng, N. Bumstead, B.F. Benkel and W.E. Briles et al., 2000. A consensus linkage map of the chicken genome. Genome Res., 10: 137-147.
PubMedDirect Link - Hassen, H., F.W.C. Neser, A. de Kock and E. van Marle-Koster, 2009. Study on the genetic diversity of native chickens in northwest Ethiopia using microsatellite markers. Afr. J. Biotechnol., 80: 1347-1353.
Direct Link - Hillel, J., M.A.M. Groenen, M. Tixier-Boichard, A.B. Korol and L. David et al., 2003. Biodiversity of 52 chicken populations assessed by microsatellite typing of DNA pools. Genet. Sel. Evol., 35: 533-557.
CrossRefPubMedDirect Link - Kalinowski, S.T., M.L. Taper and T.C. Marshall, 2007. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol., 16: 1099-1106.
CrossRefDirect Link - Kanginakudru, S., M. Metta, R.D. Jakati and J. Nagaraju, 2008. Genetic evidence from Indian red junglefowl corroborates multiple domestication of modern day chicken. BMC Evol. Biol., 8: 174-174.
PubMedDirect Link - Khatib, H., A. Darvasi, Y. Plotski and M. Soller, 1994. Determining relative microsatellite allele frequencies in pooled DNA samples. PCR Methods Appl., 4: 13-18.
PubMedDirect Link - Kimura, M. and J.F. Crow, 1964. The number of alleles that can be maintained in a finite population. Genetics, 49: 725-738.
PubMedDirect Link - Lewontin, R.C., 1972. The apportionment of human diversity. Evol. Biol., 6: 381-398.
CrossRefDirect Link - Mackey, K., A. Steinkamp and P. Chomczynski, 1998. DNA extraction from small blood volumes and the processing of cellulose blood cards for use in polymerase chain reaction. Mol. Biotechnol., 9: 1-5.
PubMedDirect Link - Moiseyeva, I.G., M.N. Romanov, A.A. Nikiforov, A.A. Sevastyanova and S.K. Semyenova, 2003. Evolutionary relationships of red junglefowl and chicken breeds. Genet. Sel Evol., 35: 403-423.
Direct Link - Mukesh, S. Sathyakumar, R.S. Kalsi, R.K. Vijh, M. Fernandes and R.P. Mandhan, 2010. Assessment of genetic diversity of red junglefowl (Gallus gallus) population in Himachal Pradesh, India. Proceedings of 5th International Galliformes Symposium Organized by World Pheasants Association, Nov. 7-14, Chiang Mai, Thailand, pp: 1-5.
- Nei, M., 1973. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA., 70: 3321-3323.
PubMedDirect Link - Niu, D., Y. Fu, J. Luo, H. Ruan, X.P. Yu, G. Chen and Y.P. Zhang, 2002. The origin and genetic diversity of chinese native chicken breeds. Biochem. Genet., 40: 163-174.
CrossRefPubMedDirect Link - Olowofeso, O., J.Y. Wang, K.Z. Xie and G.Q. Liu, 2005. Phylogenetic scenario of port-city chickens in china based on two-marker types. Int. J. Poult. Sci., 4: 206-212.
CrossRefDirect Link - Ponsuksill, S., K. Wimmers, F. Schmoll, P. Horst and K. Schellander, 1999. Comparison of multilocus DNA fingerprints and microsatellites in an estimate of genetic distance in chicken. J. Hered., 90: 656-659.
CrossRefPubMedDirect Link - Romanov, M.N. and S. Weigend, 2001. Analysis of genetic relationships between various populations of domestic and jungle fowl using microsatellite markers. Poult. Sci., 80: 1057-1063.
CrossRefDirect Link - Rosenberg, N.A., T. Burke, K. Elo, M.W. Feldman and P.J. Freidlin et al., 2001. Empirical evaluation of genetic clustering methods using multilocus genotypes from 20 chicken breeds. Genetics, 159: 699-713.
PubMedDirect Link - Tadano, R., M. Nishibori, N. Nagasaka and M. Tsudzuki, 2007. Assessing genetic diversity and population structure for commercial chicken lines based on forty microsatellite analyses. Poult. Sci., 86: 2301-2308.
Direct Link - Vanhala, T., M. Tuiskula-Haavisto, K. Elo, J. Vilkki and A. Maki-Tanila, 1998. Evaluation of genetic variability and genetic distances between eight chicken lines using microsatellite markers. Poult. Sci., 77: 783-790.
CrossRefPubMedDirect Link - Bao, W.B., G.H. Chen, X.S. Wu, Q. Xu, S.L. Wu, J.T. Shu and S. Weigend, 2007. Genetic diversity of red jungle fowl in China (Gallus gallus spadiceus) and red jungle fowl (Gallus gallus gallus) in Thailand. Yi Chuan, 29: 587-592.
PubMed - Ya-Bo, Y., W. Jin-Yu, D.M. Mekki, T. Qing-Ping and L. Hui-Fang et al., 2006. Evaluation of genetic diversity and genetic distance between twelve Chinese indigenous chicken breeds based on microsatellite markers. Int. J. Poult. Sci., 5: 550-556.
CrossRefDirect Link