
Juan Gualberto Colli
Unidad de Bioquirnica y Biologia Molecular de Plantas, Centro de Investigaci6n Cientifica de Yucatan, Calle 43 No. 130, CoL Chubwmi de Hidalgo, CP 97200, Merida, Yucatim, Mexico
Diana Guillen
Unidad de Bioquirnica y Biologia Molecular de Plantas, Centro de Investigaci6n Cientifica de Yucatan, Calle 43 No. 130, CoL Chubwmi de Hidalgo, CP 97200, Merida, Yucatim, Mexico
Luis Carlos Rodriguez
Unidad de Biotecnologia, Centro de Investigacion Cientifica de Yucatan, Calle 43 No. 130, CoL Chubwmi de Hidalgo, CP 97200, Merida, Yucatim, Mexico
Nadine Mobious
Unidad de Bioquirnica y Biologia Molecular de Plantas, Centro de Investigaci6n Cientifica de Yucatan, Calle 43 No. 130, CoL Chubwmi de Hidalgo, CP 97200, Merida, Yucatim, Mexico
Enrique Castano
Unidad de Bioquirnica y Biologia Molecular de Plantas, Centro de Investigaci6n Cientifica de Yucatan, Calle 43 No. 130, CoL Chubwmi de Hidalgo, CP 97200, Merida, Yucatim, Mexico
Asian Journal of Biochemistry
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 75-83







ABSTRACT
Transcriptional regulation depends on the appropriate set of positive and negative regulating signals in order to provide the correct gene expression. Studies in eukaryotic gene expression over the last few years have shown that SRBs/Mediator complex is involved in the proper activation of a variety of genes, while NC2 has been shown to be a general repressor of transcription. NC2 is a conserved transcripcional regulator that was originally identified as inhibitor of basal transcription. Here Our initial results show that the repressor NC2 can interact directly with the Mediator in HeLa cells, this interaction provides a new set of interactions between positive and negative cofactors. Furthermore NC2 depletion from HeLa nuclear extract decrease activated transcription from the HIV–LTR1 promoter in vitro but addition of a complex NC2 fraction restores activated transcription. Further dissection showed that NC2 interacts directly with MED21 a subunit of the Mediator complex. The results shown may provide clues to the controversial role that NC2 has on active promoters.
PDF Abstract XML References Citation
How to cite this article
Juan Gualberto Colli, Diana Guillen, Luis Carlos Rodriguez, Nadine Mobious and Enrique Castano, 2006. Direct Interaction Between NC2 and a Subunit of the SRBs/mediator Co–activator. Asian Journal of Biochemistry, 1: 75-83.
DOI: 10.3923/ajb.2006.75.83
URL: https://scialert.net/abstract/?doi=ajb.2006.75.83
DOI: 10.3923/ajb.2006.75.83
URL: https://scialert.net/abstract/?doi=ajb.2006.75.83
INTRODUCTION
Transcriptional activation and repression of class II genes has been the subject of intense study since the last three decades. During the early 90’s, a repressing activity was found in HeL a nuclear extract that could selectively reduce basal transcription but did not affect the activated transcription. Consequently, this activity was named Negative Cofactor 2 (NC2) (Kim et al., 1995). Soon after, protein Dr1 was found to be a general transcription repressor, particularly when it is associated to its partner DRAP1. In addition, NC2 was cloned and found to be the same as Dr1/Drap1 (Goppelt et al., 1996). The recombinant proteins have been found to repress transcription in vitro by interacting with TBP thereby preventing TFIIB from forming a DNA-TBP-TFIIB complex. The result is the inhibition of the preinitiation complex formation while maintaining a DNA-TBP-NC2 complex (Goppelt et al., 1996). TFIIA competes with NC2 for TBP and allows TFIIB to bind, which promotes binding from the rest of general transcription factors and RNA Pol II to the promoter and for transcription initiation to occur. In the early 90’s several positive transcriptional activities were found, one particularly known as the Positive Cofactor 2 (Kretzschmar et al., 1994). As it is known to stimulate GAL4-AH in an in vitro transcription assay, several studies have shown that the polypeptides involved in this activity belong to what has now been called the Mediator complex, depending on the conditions in which it has been purified. It has yielded different complexes such as TRAPS, CRSP, ARC, DRIP, Mediator, etc. For reviews, see several published articles (Kretzschmar et al., 1994, Ito et al., 2001, Naar et al., 2001, Bikle et al., 2003, Bjorklund and Gustafsson, 2004). It is still not know if there are many complexes that share the same subunits or that only one function as a general cofactor for different activators with small differences, depending on the purification protocol involved to obtain them. It is accepted that these sets of polypeptides interact with the basal transcription machinery and the activators, providing the link between them and therefore a higher control and yield of transcription.
In yeast, activators have been shown to bind directly to the SRB-Mediator complex (Davis et al., 2002) and genetic and biochemical studies have identified the SRB4 as a target of the well-studied acidic activator Gal4 (Kamada et al., 2001). Temperature-sensitive mutations SRB4 subunit can produce a rapid, general shutdown of mRNA synthesis, demonstrating that SRB4 is required for expression of most protein-encoding genes (Christova and Oelgeschlager, 2002). Because essentially all of the SRB proteins are tightly associated with the holoenzyme in yeast cells, the SRB-containing holoenzyme likely functions in transcription initiation at most class II promoters in vivo. The mutant genes that compensated for the absence of SUC2 activator function included genes called BUR genes (Bypass UAS Requirement). The bur6 mutant allele was found to be a identical to NCB1(NC2a) indicates that a loss in NC2 function can compensate for the loss of an activator. These results point out to a delicate balance between positive and negative regulators of transcription which operate through the Ydr1-Bur6 (yeast NC2) repressor complex (Davis et al., 2002).
Over the years the simple model of repression via NC2 has been challenged, as new data have suggested that NC2 may be found in active promoters. Such has been the case in HeLa cells where it has been found in active promoters (Kim et al., 1995) in yeast cell analysis in genetic and biochemical studies (Creton et al., 2002, Denko et al., 2003). In Drosophila´s TATAless promoters was shown to be required as a general transcription factor (Willy et al., 2000). We previously found NC2 to interact directly or indirectly with RNA pol IIO (Castano et al., 2000).
Recent work has shown DR1 disassociated from DRAP1, depending on the conditions of cell growth and tissue (Ikeda et al., 1998, Iratni et al., 2002). Either could be due to different binding partners yet to be found, currently is known the interaction with AREB6 (Ikeda et al., 1998) and FoxH1 (Iratni et al., 2002). Until now, it has not been shown a clear physical connection between NC2 and the positive cofactors. These previous studies provide a correlation between the presences of both NC2 subunits and other proteins, but do not address the role of NC2 during activation. The question as to how does NC2 stimulates transcription remains unknown.
In this study we have report a direct interaction between NC2 and MED21 (previously known as hSRB7). The interaction was verified by NC2 co-depletions in nuclear extract as well as with pure recombinant proteins. Furthermore we found that upon removal of NC2 from HeLa nuclear extract the level of activated transcription in a cell free system is significantly reduced but addition of chromatographic fractions containing NC2 and the Mediator can restore transcriptional activation. This information together with the novel interaction between NC2 and MED21 suggest a balance of positive and negative transcriptional responses.
MATERIALS AND METHODS
Vectors
The vector pNC2 α-6HIS and pNC2β-6HIS have previously been described. The vector pNC2α-M2 was carried out by subcloning NC2α with NdeI and Bam HI into pET11d-flag. The final co-expressing vector was carried out by subcloning NC2α-M2 with Bgl II and Hind III in to pNC2β-6HIS, in order to generate pNC2β-6HIS/NC2α-M2. Promoter Constructs. The pHIV-111/80 containing SP1 activation sites and pHIV-34/60 containing the core promoter has been described elsewhere (Castano et al., 2000).
NC2 Expression the vectors were transformed in LysE bacteria and then allowed to grow to an OD of 0.1. Next, IPTG was added and incubated at 30 degrees (C) for 6 h Bacterially expressed M2: NC2α- 6His:NC2β and 6His:hSRB7 were purified on Ni2+- nitrilotriacetic acid resin under non denaturing conditions. M2: NC2α- 6His:NC2β were further purified over an M2 resin column in BC buffer (20 mM TrisAHCl, pH 7.9 at 4°C/20% glycerol/0.2 mM EDTA, pH 8.0/10 mM β-mercaptoethanol/0.5 mM PMSF) containing 0.1 M KCl.
Protein Extraction
Extracted and purified proteins were stored in BC buffer containing 0.1 M KCl. 15 L of HeLa cells were used to prepare the nuclear extract (HeLa NE) as described previously (Dignam et al., 1983) HeLa NE lacking NC2 was prepared by immunoaffinity chromatography on protein A-Sepharose CL4B (Amersham Pharmacia) covalently crosslinked to polyclonal anti-NC2β antibody. Immunodepletions were carried out in BC buffer containing 0.5 M KCl and 0.1% NP-4010 mL of HeLa Nuclear Extract was equilibrated with BC buffer containing 300 mM KCl. HeLa NE was clarified by centrifugation at 10,000 g for 20 min and loaded into a Phosphocellulose P11 (Whatman), previously equilibrated with the BC 300. Once the entire volume is loaded, the column is washed with 3-5 column volumes (CVs) of BC 300. Elute the column with 3 CVs of BC buffer with 500 mM KCl. Twice dialyzed the eluted fraction with BC 100 was applied to the DEAEff column previously equilibrated in BC 100. Collect protein fractions. Collected flow through fractions is directly pass over a CM sepharose column (Amersham Pharmacia) previously equilibrated with BC 100. CM sepharose column was washed 5 CVs of BC100. Elution was carried out with 5 CVs of BC 400. Pool fractions based on Abs280 nm, colorimetric protein assay,
Immunoprecipitations
They were conducted as described earlier (Creton et al., 2002). In brief the protein A beads were crosslink with anti NC2β , followed by being incubated at 4 degrees for 2 h with purified NC2. Next, removal of non-bound NC2 was completed by washing three times the pellet with BC buffer containing 0.3 M KCl and 0.1% NP-40, the NC2 beads were then incubated with purified MED21 for 2 h at 4 degrees. The immunoprecipitations were carried out in BC buffer containing 0.3 M KCl and 0.1% NP-40. Moreover, size exclusion chromatography was carried out on FPLC with Superose 12 from Amersham Pharmacia, in a BC buffer containing 0.3 M KCl.
Western Blot Analysis
Untreated HeLa NE and NC2β-depleted HeLa NE (NE[ΔNC2β]). Equal amounts of NE protein (30 μg) and 10 μL of anti-NC2β column eluate were subjected to 10% SDS/PAGE, transferred to nitrocellulose and analyzed with antibodies to the proteins indicated.
Farwestern
Hundred micrograms of rNC2 were label with 32P gATP in 20 units of casein Kinase II for 1 h at 30 degress. Labelled rNC2 was purified by Ni2+- nitrilotriacetic acid resin. The labelled NC2 was used to probe the re-natured blot from the eluted NC2 of the immunoaffinity chromatography. Re- naturation of the blot was carried out by incubating the membrane blot in 8 M urea TBST followed by a 4 M urea TBST and a subsequent 2 M urea TBST before a final wash in TBST, all incubations were carried out for 2 h at room temperature. After the membrane had been re-natured incubations with the label NC2 were carried out at 4 degrees in TBST for 4 h after several washes with TBST and Autoradiography analysis. The same membrane was also used for western blot analysis with anti-MED21.
In Vitro Transcription
Standard 60-min in vitro transcription reactions were performed as described previously (Castano et al., 2000). In brief a reaction mixture containing either nuclear extracts or purified transcription factors were mix with 100 ng of pTOG5 ML in the presence of 0.5 mM NTP, 5 mM MgCl, 5 mM DTT, in 20 mM HEPES KOH pH 8.4. in 20 μL final volume. The transcription reactions were incubated at 30°C for 1 h. A control label DNA was added to the mixture. Products were extracted in phenol, ethanol-precipitated and used for primer extension reactions utilizing 32P label primer as previously published (Ikeda et al., 2002).
RESULTS
NC2 has been shown to be a general transcriptional repressor of class II genes and its biochemistry has been well-characterized. The structure and interactions with TBP have been formally proven by x-ray crystallography (Kamada et al., 2001). However, in order to achieve a further understanding of the function of this repressor, an immunodepletion from HeL a, nuclear extract with anti-NC2β was carried out.
Surprisingly our experiments show that members of the SRB/Mediator complex co-precipitated selectively with NC2 as shown in Fig. 1. The lack of precipitation of general transcriptions factors suggested a high specificity of the assay although several nonspecific proteins do bind to the resin. Together with the tied binding of the anti NC2 antibody make the system unsuitable for further protein purification. Nevertheless many possible interacting partners could be analyzed from this system. Four antibodies directed against the SRB/Med complex showed a positive signal in the co-elution with NC2, however only MED21 showed around a 50% co-depletion in the NC2 depleted extract, (Fig. 1). Moreover as can be observed NC2 does not interact with generals factors of transcription such as TFIIB y TFIIH, etc. Pointing to a specific interaction between the MED21 and NC2, the lack of depletion of other Mediator components can be explain by the great amount MED21 found in the cells that is not bound to the other subunits.
In order to evaluate the functional effect of NC2 depleted from nuclear extract. in vitro transcription reactions where carried out. As can be seen in Fig. 2A a 89% decrease in activated transcription is observed when NC2β is depleted from nuclear extract, however the addition of the chromatographic fraction P11:05 contiaing NC2 and Mediator complex can completely restore transcriptional activation this is not the case for other chromatographic fractions or recombinant NC2. This suggest that NC2 is insufficient for the increase in transcriptional activation, indicating missing elements that were depleted from the nuclear extract. Different resins have been used in vane to try to further purify the activity as shown in Fig. 2B and 2C.
 | |
| Fig. 1: | Immunoblot analysis of HeLa NE, NC2β depleted HeLa NE and the eluate from NC2β Immunocolumn. Antibodies again NC2α, NC2β, hSUR2, hMED7, TFIIB, CDK8, TFIIH, TAF100, MED21 and were used to probe the membranes for their presence. Protein molecular weight markers were used for all cases (pm) |
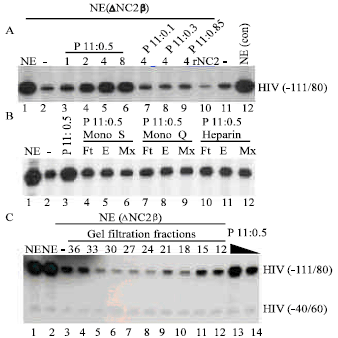 | |
| Fig. 2: | NC2 depletions decrease activated transcription in vitro. A. in vitro transcription With HeLa nuclear extract (NE), preimmune depleted nuclear extract (NE control), NC2 depleated nuclear extract (αNC2β NE), Phosphocellulose 11 fractions from nuclear extract either at 0.1, 0.3, 0.5 or 0.85 M KCL correspond to P11:0.1, P11:0.3, P11:0.5, P11:0.85 respectively. B in vitro transcription testing eluate fractions, flowthrow or mixing experiments from either Mono S, Mono Q or Heparin chromatographic resins utilizing P11:0.5 as the material source. C. in vitro transcription testing the effect of size exclusion chromatography of the P11:0.5 fraction |
 | |
| Fig. 3: | Farwestern blot analysis. A, Autoradiograph of a 12% PAGE of Casein kinase II labelled and purified rNC2. B, Farwester blot analysis of NC2 eluate from the immunopreciptate with anti-NC2β from HeLa NE, after renaturation. As described in materials and methods C. Immunoblot analysis from the membrane blot was carried out with anti-hSRB7 |
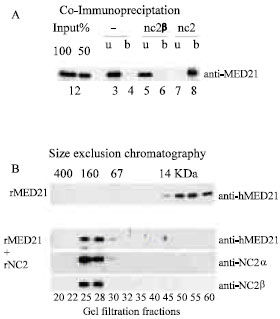 | |
| Fig. 4: | Direct interaction between hSRB7 and NC2. A, Co-Immunoprecipitation of MED21 with NC2. Cross link anti-NC2β to agarose beads was used as in the absence, or presence of NC2β or NC2 complex. After washing the excess nonbound NC2 the MED21 was allowed to interact as described in Materials and Methods. B, Superose 12 gel filtration chromatography was carried out. Purified hSRB7 MED21 showed a low mobility under 14 KDa in the absence of other proteins and high mobility of 160 KDa when incubated in the presence of purified rNC2 |
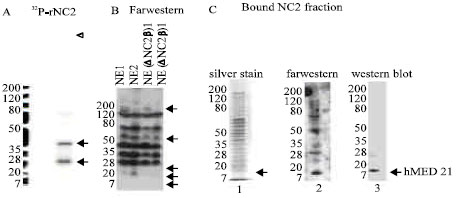
In order to define what other factor may be interacting with NC2, we labeled the NC2 complex with 32Pγ ATP and casein kinase II, Fig. 3A shows the autoradiography both labeled rNC2 subunits. A farwestern approach was carried on two different nuclear extracts to identify those proteins that interact with NC2. Several bands appeared but the main bands where found at 70, 50, 35, 30, 25 and 19 Kda as displayed in Fig. 3B. was of particular interest as it showed a consistent decrease of a 19 KDa band in depleted NC2β extracts. Futheremore, a farwestern carried out on the NC2 eluate shows an interacting protein of 19 KD as seen in lane 2 of Fig. 3C. Many of the bands are not due to nonspecific binding as the overall protein amount found in this region is low as seen in Fig. 3C lane 1. Lane 3 from Fig. 3C show an in munoblot analysis of the farwestern with anti-hMED21 showing that the 19 KDa band match the signal from the farwestern from lane 2. We thus tested the direct interaction of NC2 with MED21 since it showed a higher yield of co- precipitation from the in muno-de pletion assay and had a molecular weight matching the 19 KDa interacting protein from the farwestern. To test for a possible interaction In munoprecipitations were carried out with the purified proteins which showed a greater than 90% interaction of MED21 with NC2 complex but no binding was observed with NC2β or the antibody bound to the beads as seen on Fig. 4A.
Finally, to further prove the interaction and avoid the use of antibodies in the assay, a gel filtration column was used to separate the non interacting proteins. A change was observe in the mobility of all detectable MED21 when it is incubated with NC2 to coincide with the peak of elution of NC2, thereby showing a physical interaction of MED21 with NC2 as shown in Fig. 4B.
DISCUSSION
NC2 (Dr1/Drap1) has long been known to be one of the main general transcriptional repressors in eukaryotic cells (Goppelt et al., 1996, Yeung et al., 1997). The interaction between NC2 and TBP has been demonstrated in vitro, showing its role as a repressor by preventing TBP interaction with TFIIB thereby blocking the formation of the pre-initiation complex (Geisberg et al., 2001). Furthermore, plants NC2 shows an inverted role in the transcriptional repression of the two subunits (Song et al., 2002). However, resent literature has suggested that NC2 can be found dissociated and interacts with other polypeptides like AREB6 and FoxH1 (Ikeda et al., 1998; Iratni et al., 2002). One possibility is that the histone fold motive common to a few polypeptides may help them being interchangeable depending on the cellular response which modulates the transcriptional response.
NC2 has been shown in Drosophila TATAless promoters as a general transcription factor (Willy et al., 2000) it still remains uncertain what is the effect of NC2 in human TATAless promoters. Moreover it has also been reported a dual activity of NC2 in yeast experiments (Gadbois et al., 2003). It has been shown that in yeast cells the loss of NC2 function can compensate for a defect in the SRB4 component of the holoenzyme, together with previous evidence that SRB4 functions positively in activation at a global level in class II promoters it may reflect that NC2 interact with the holoenzyme in a balance activation and repression (Gadbois et al., 2003). Furthermore NC2 alpha has been shown to interact with BTAF1 and this may lead to stimulation in its ATP-dependent association with TBP, thereby increasing its affinity to DNA (Klejman et al., 2004). Present results show that a different member of the SRB/mediator complex (MED21) physically interacts with NC2 in human extracts and purified proteins as seen in Fig. 1 and 4.
However, there is little conservation on several portions of the sequence between human, yeast and Drosophila NC2. The function of NC2 in our in vitro transcription experiments show a positive role for this repressor, since its depletion from HeLa nuclear extract results in a ten fold decrease of activated transcription Fig. 2. rNC2 was incapable of rescuing transcription activation and resulted in a further decrease of transcription, consistent with other publish reports (Kim et al., 1995) but suggesting that other factors may be lost during the NC2 depletion. The addition of P11:0.5 fractions obtained from HeLa nuclear extract can rescue the lost transcriptional activity of the NC2 depleated extract. This fraction contains the majority of NC2 from nuclear extract, but its purification over resins does not yield further purification of the activity, surprisingly the activity was lost over a size exclusion chromatography, which points out to either to protein modifications (dephosphorylation, methylation, sumorylation, etc) which may occur during the purification procedure that renders the complex inactive or dissociates it from other proteins. Also can be explain if weak binding of the large complex Mediator dissociates some components during the affinity purification rendering the left complexes inactive. Further advances in protein purification techniques as well as the creation of tag cell lines may be need it to answer this problem.
The precipitate from the immunodepletion and the farwestern blot show that NC2 can interact with several proteins (Fig. 3). To confirm this finding, further work is necessary to elucidate all of the partners and condition under which this protein may be having a biological response. However the finding by Farwesterns of a 19 KDa protein that correspond by western blot to MED21 and the CO-IP and Gel filtration suggest a strong interaction, current yeast data suggest that SRB4 plays a regulatory role with yeast NC2 (Gadbois et al., 2003).
In summary, we found in this research that NC2 and MED21 interact and help modulate the level of transcription by the SRBs/Mediator complex, or it may interact with some of the free portion of cellular MED21, which role is still to be defined.
ACKNOWLEDGMENTS
We thank Arnold Berk for the anti-hSUR2 antibody and Yi Wei Jiang for anti-hMED7 antibody. We wish to thank Angela F. Ku for her technical help, T. Oelgeshlager group for their discussions and Luis Joel Figeroa from Accesolab for the contribution of reagents for this study. Dr. Castano is partially funded by CONACYT grant 39731-z.
REFERENCES
- Bikle, D.D., C.L. Tu, Z. Xie and Y. Oda, 2003. Vitamin D regulated keratinocyte differentiation: Role of coactivators. J. Cell Biochem., 88: 290-295.
Direct Link - Castano, E., P. Gross, Z. Wang, R.G. Roeder and T. Oelgeschlager, 2000. The C terminal domain phosphorylated IIO form of RNA polymerase II is associated with the transcription repressor NC2 (Dr1/DRAP1) and is required for transcription activation in human nuclear extracts. Proc. Natl. Acad. Sci. USA., 97: 7184-7189.
PubMed - Christova, R. and T. Oelgeschlager, 2002. Association of human TFIID promoter complexes with silenced mitotic chromatin in vivo Natl. Cell Biol., 4: 79-82.
PubMed - Creton, S., J.Q. Svejstrup and M.A. Collart, 2002. The NC2 α and β subunits play different roles In vivo. Genes Dev., 16: 3265-3276.
CrossRef - Davis, J.A., Y. Takagi, R.D. Kornberg and F.A. Asturias, 2002. Structure of the yeast RNA polymerase II holoenzyme Mediator conformation and polymerase interaction. Mol. Cell, 10: 409-415.
PubMed - Denko, N., K.W. Dollries, A.B. Johnson, E. Hammond, C.M. Chiang and M.C. Barton, 2003. Hypoxia actively represses transcription by inducing negative cofactor 2 (Dr1/DrAP1) and blocking preinitiation complex assembly. J. Biol. Chem., 278: 5744-5749.
CrossRef - Dignam, J.D., P.L. Martin, B.S. Shastry and R.G. Roeder, 1983. Eukaryotic gene transcription with purified components. Methods Enzymol., 101: 582-598.
Direct Link - Gadbois, E.L., D.M. Chao, J.C. Reese, M.R. Green and R.A. Young, 1997. Functional antagonism between RNA polymerase II holoenzyme and global negative regulator NC2 in vivo. Proc. Natl. Acad. Sci. USA., 94: 3145-3150.
Direct Link - Geisberg, J.V., F.C. Holstege, R.A. Young and K.Y. Struhl, 2001. Yeast NC2 associates with the RNA polymerase II preinitiation complex and selectively affects transcription in vivo. Mol. Cell Biol., 21: 2736-2742.
CrossRef - Goppelt, A., G. Stelzer, F. Lottspeich and M. Meisterernst, 1996. A mechanism for repression of class II gene transcription through specific binding of NC2 to TBP-promoter complexes via heterodimeric histone fold domains. EMBO J., 15: 3105-3116.
Direct Link - Ikeda, K., J.P. Halle, G. Stelzer, M. Meisterernst and K. Kawakami, 1998. Involvement of negative cofactor NC2 in active repression by zinc finger homeodomain transcription factor AREB6. Mol. Cell Biol., 18: 10-18.
Direct Link - Iratni, R., Y.T. Yan, C. Chen, J. Ding and M.M. Shen et al., 2002. Inhibition of excess nodal signaling during mouse gastrulation by the transcriptional corepressor DRAP1. Science, 298: 1996-1999.
PubMed - Ito, M. and R.G. Roeder, 2001. The TRAP/SMCC/Mediator complex and thyroid hormone Trends Endocrinol Metab., 12: 127-134.
PubMed - Kamada, K., F. Shu, H. Chen, S. Malik and S.K. Burley et al., 2001. Crystal structure of negative cofactor 2 recognizing the TBP-DNA transcription complex. Cell, 106: 71-81.
Direct Link - Kim, T.K., Y. Zhao, H. Ge, R. Bernstein and R.G. Roeder, 1995. TATA binding protein residues implicated in a functional interplay between negative cofactor NC2 (Dr1) and general factors TFIIA and TFIIB. J. Biol. Chem., 270: 10976-10981.
Direct Link - Klejman, M.P., L.A. Pereira, H.J. Van Zeeburg, S. Gilfillan, M. Meisterernst and H.T. Timmers, 2004. NC2alpha interacts with BTAF1 and stimulates its ATP-dependent association with TATA-binding protein. Mol. Cell Biol., 24: 10072-10082.
Direct Link - Kretzschmar, M., G. Stelzer, R.G. Roeder and M. Meisterernst, 1994. RNA polymerase II cofactor PC2 facilitates activation of transcription by GAL4 AH in vitro. Mol. Cell Biol., 14: 3927-3937.
PubMed - Naar, A.M., B.D. Lemon and R. Tjian, 2001. Transcriptional coactivator complexes. Annu. Rev. Biochem., 70: 475-501.
PubMedDirect Link - Prelich, G., 2002. Saccharomyces cerevisiae BUR6 encodes a DRAP1/NC2alpha homolog that has both positive and negative roles in transcription in vivo. Mol. Cell. Biol., 17: 2057-2065.
CrossRefDirect Link - Song, W., H. Solimeo, R.A. Rupert, N.S. Yadav and Q. Zhu, 2002. Functional dissection of a rice Dr1/DrAp1 transcriptional repression complex. Plant Cell., 14: 181-915.
CrossRef - Willy, P.J., R. Kobayashi and J.T. Kadonaga, 2000. A basal transcription factor that activates or represses transcription. Science, 290: 982-984.
CrossRefDirect Link - Yeung, K., S. Kim and D. Reinberg, 1997. Functional dissection of a human Dr1-DRAP1 repressor complex. Mol. Cell. Biol., 17: 36-45.
Direct Link