
N.M. Azuadi
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
S.S. Siraj
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
S.K. Daud
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
A. Christianus
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
S.A. Harmin
Centre of Land and Aquatic Biotechnology, Faculty of Science and Biotechnology, Universiti Industri Selangor, 45600, Bestari Jaya, Selangor, Malaysia
S. Sungan
Department of Agriculture, Indigenous Fisheries Research and Production Centre, Tarat, Sarawak, Malaysia
R. Britin
Department of Agriculture, Indigenous Fisheries Research and Production Centre, Tarat, Sarawak, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 6 | Page No.: 761-774







ABSTRACT
The study was conducted to observed the embryonic development, morphology of eggs, newly hatched larve and post larval of the filial 1 (F1) generation of Malaysian mahseer, Tor tambroides using hatchery-reared specimens. The first cleavage occurred at 2 h after fertilization (HAF), epiboly began at 11 HAF, the embryonic body was formed at 22 HAF and hatched at 76 HAF. The mean total length of newly hatched larvae was 7.44±0.06 mm and almost has similar external features to other Tor species. The mouth and foregut opened at 7th Day after Hatching (DAH) with the mean total length of 12.8±0.11 mm. Tor tambroides reached 13.2±0.15 cm of total length and 31±0.53 g of body weight at 10 months of age. The induced ovulation technique using newly developed commercial synthetic hormone, Ovatide have paved the way for further development and improvement of culture production of the Malaysian mahseer.
PDF Abstract XML References Citation
Received: March 15, 2013;
Accepted: August 21, 2013;
Published: September 25, 2013
How to cite this article
N.M. Azuadi, S.S. Siraj, S.K. Daud, A. Christianus, S.A. Harmin, S. Sungan and R. Britin, 2013. Induced Ovulation, Embryonic and Larval Development of Malaysian Mahseer, Tor tambroides (Bleeker, 1854) in Captivity. Asian Journal of Animal and Veterinary Advances, 8: 761-774.
DOI: 10.3923/ajava.2013.761.774
URL: https://scialert.net/abstract/?doi=ajava.2013.761.774
DOI: 10.3923/ajava.2013.761.774
URL: https://scialert.net/abstract/?doi=ajava.2013.761.774
INTRODUCTION
The Malaysian mahseer or Tor tambroides, belonging to family Cyprinidae, is an attractive and important freshwater species for recreational fisheries and aquaculture in Malaysia. It is known for its delicacy and firm flesh as food and aggressive as sport fish. It is one of the highly priced freshwater fish species as its demand exceeds the supply. The market price in Malaysia ranges from RM 370-700 (USD 117-228.12) per kg.
Spawning in mahseer takes place after rainy period suggesting that monsoon season which normally occurred from December to March influences the spawning (Ingram et al., 2007). Tor tambroides is asynchronous spawners and capable of spawning several times per year (Ingram et al., 2007) but the highest spawning activities occur during monsoon season. Due to this restrictive reproductive nature, the fish cannot ovulate or spawn all year around. Thus, introduction of sustained release of gonadotropin-releasing hormone analogue (GnRHa) such as Ovaplant, prior to hormone treatment will advance the final oocyte maturation in T. tambroides and improves the likelihood of ovulation induction in captivity (Ingram et al., 2005).
In Malaysia, the first successful induced ovulation recorded in Tor tambroides was by Ingram et al. (2005). They used pond-held broodstocks (initially wild-caught) for artificial propagation. Breeding performances of the breeders, however, was still low and lead to low quality fry. Since the culture of T. tambroides is closely dependent on the availability of their fry, the present collection of fry from nature cannot satisfy the demand. In addition, mature T. tambroides breeders are not always available from the natural habitat because of the declining natural populations due to habitat degradation and overfishing. Thus, it is desirable to develop and establish new techniques for artificial propagation for this fish. Ovulation induction by hormonal treatment is one of the ways to produce fry for stocking, together with a supply of good quality mature female broodstock to ensure continuous mass production of juveniles.
Previous studies showed that T. tambroides do not undergo final oocyte maturation, ovulation or spawning in captivity (Azuadi et al., 2011). Without exogenous hormone stimulation, ova will undergo atresia and degenerate and subsequently will not proceed to final oocyte maturation and ovulation (Mousa, 2010). There are a number of hormones available in market. Ovatide, a newly introduced commercial hormone in Malaysia, is cheaper and less viscous than Ovaprim. In India, Ovatide has been used effectively in inducing final oocyte maturation and spawning in other cyprinids, such as Indian major carps Cirrhina mrigala and Labeo rohita (Dhawan and Kaur, 2004). To date there are no results or reports on effectivenss of Ovatide to induce ovulation of T. tambroides. Although, Ovaprim is the most successful hormone to induce ovulation of wild caught T. tambroides but the result was not too promising (Ingram et al., 2005). Hence, the use of Ovatide is preferred over Ovaprim in this study.
The aims of this study were to use Ovatide for inducing ovulation in filial 1 generation of T. tambroides breeders in captivity and to describe the ovulation, egg, larval and post larval developments.
MATERIALS AND METHODS
Broodstock management: Sexually matured filial 1 (F1) generation of female Tor tambroides were stocked indoor in 20 m3 concrete tanks using Recirculating Aquaculture System (RAS) at Lu Thian Tack (LTT) Aquaculture Farm, Asajaya, Sarawak. The fish were obtained from Tarat Indigenous Fisheries Production and Research Center (IFPRC), Serian, Sarawak. Water was maintained at the temperature ranged from 25.0-27.5°C, pH ranged from 7.21-8.02 and dissolved oxygen concentration above 6.0 mg L-1. In the morning, breeders were fed with sinking homemade pelleted diet (vitamin C in combination with illipenut (Engkabang) fruit, emperang fish, wheat flour and squid oil) at 5% of the body weight. The feed contained 52.6% crude protein, 17.6% crude lipid, 1.8% crude fibre, 5.3% carbohydrate, 12.7% ash and 4770.5 k cal g-1 energy. In the evening, the fish were fed with commercial Tilapia pellet (Cargill, Malaysia) containing 16% protein, 4% fat and 6% crude fiber. Matured Tor tambroides broodstocks (at least 4 years old) with total weight ranging from 2.50-3.93 kg and total length ranging from 50.8-63.9 cm were used. A total of 14 females and 7 males were selected for this study. Males were identified with running ripe milt and females with slightly distended abdominal condition (Azuadi et al., 2011).
Acclimation and induction of ovulation: The present study was carried out during the natural spawning season of Tor tambroides at (LTT) Aquaculture Farm, Asajaya, Sarawak from November 2010-February 2011. The fish were anesthetized in a solution of tricaine methanesulfonate MS222 (100 mg L-1) (Syndel International Inc., Qualicum Beach, BC, Canada) before handling (Ingram et al., 2005). Selected broodstocks were acclimated in 15000-l rectangular cement tanks. The tanks were equipped with recirculation system with running ozonated fresh water (20%) and aeration.
Six weeks prior to induction, matured F1 generation breeders were implanted with Ovaplant (75 or 150 μg), Syndel International Inc., Qualicum Beach, BC, Canada. Ovatide at 0.5 mL kg-1 BW dosage was injected intramuscularly adjacent to the dorsal fin. No hormonal injection was needed for the males since there was no problem of getting running milt at anytime. Twenty four h after administration, ovulatory response was evaluated by applying gentle pressure on the female’s abdomen. Egg volume was recorded to estimate the total egg production and stripping fecundity. Matured males with running ripe milt were chosen and stripped into the jar containing eggs for fertilization using dry method of Joshi et al. (2002).
Collected data were analysed based on the (1) No. of stripped eggs, (2) Fertilization rate, (3) Hatching rate, (4) Survival rate, (5) Normal hatched larvae and (6) Deformed hatched larvae. Number of ovulated eggs was estimated by multiplying of number of egg mL-1 with total volume of eggs for each female. Fecundity was calculated by dividing total number of eggs by female body weight. Fertilization and hatching rate was determined by taken 200 egg samples from each breeder with 3 replicates. Survival, normal and deformed larvae rates were determined by randomly taken three replicates of 200 larvae for each breeder. Deformed larvae were recorded to calculate normal larvae production. Deformed larvae were determined by observing those larvae having abnormal head, bent tail, bent notochord and bloated body. Fertilization, hatching, survival, normal and deformed rates were calculated by using the following formulae:
 |
Eggs incubation and embryonic development: Fifty fertilized eggs were randomly taken to measure egg diameters. Viable fertilized eggs from the different female breeders were stocked in plastic mesh tray incubators (volume of 400 mL with mesh size, 1000 μm) at 80 eggs mL-1. The incubation trays were placed on water surface in 1000 L rectangular fiberglass tanks. During the experiment, the flow rate of water into the tanks was 1-2 L ha-1, treated with UV and filtered. In order to determine the embryonic development, 50 eggs were taken from each incubation tray every 1 h until the blastula stage. Photographs of egg development were taken using stereomicroscope, model Motic SMZ-168 (2x magnification, 34.5 mm working distance).
Larval and post larval development: For larval development, 50 samples were taken for observation and measurement from the day of hatching until 45 Days after Hatching (DAH). Total length measurement of each sample was taken under stereomicroscope by using vernier caliper. At 45 DAH, the larvae were transferred into separate happanets in cement tanks. Each happa net contained 200 larvae with a total of 5 replicates for the experiment. Ten samples of fingerlings at 10 months of age were taken from each happa net for measurement of total length and weight. Photographs were taken using Canon digital camera at 90 DAH, 200 DAH and 10 month DAH.
Feeding regime: At 7 DAH, the larvae were given live feed, Artemia nauplii, until 14 DAH. At 14 DAH, the feed was changed into mesh artificial feed containing fishmeal and pellet (32% protein, Otohime Japan). At 90 DAH, the larvae were introduced with small granule pellet (50% protein, Otohime Japan) until 200 DAH. At 200 DAH, the larvae were fed with 2 mm pellet containing 32% protein (Otohime Japan). After 10 months of hatching, the fish were given 3 mm pellet with 32% protein content (Otohime Japan).
Egg and larval development analysis: The embryonic development of filial 1 (F1) generation of Malaysian mahseer, Tor tambroides was described based on terminology and staging that were proposed for the common development of teleosts by Saka et al. (2001), Kamaci et al. (2005) and Mousa (2010). Furthermore, to clarify the similarities and differences in embryonic development among species, results were compared with previous research into Tor tambroides, Tor duronensis, Tor putitora and Tor kudree.
RESULTS
Induced ovulation: Eighteen female broodstocks showed signs of maturation by having soft and rounded belly with protruding and swollen papillae after 6 week of implantation. High rate (78%) of ovulation success was achieved in Tor tambroides females administered with 0.5 mL kg-1 of b.wt. of Ovatide. All females (N = 18) observed underwent ovulation at 23-25 h post administration. The number of stripped eggs, the fertilization, hatching and survival rates in different females are shown in Table 1. The number of ovulated eggs ranged from 652 and 3900 with the mean value of 1599±256. The rates of fertilization, hatching and survival ranged from 80.1-99.9%, 55.0-96.4% and 60.0-91.4% with the mean values of 91.2±2.6, 80.5±2.9 and 79.7±2.4, respectively. High rate of normal and low rate of deformed hatched larvae were obtained in this experiment with a mean value of 81.95±2.2 and 18.86±2.2, respectively.
| Table 1: | Breeding performance of F1 Tor tambroides induced with 0.5 mL kg-1 ovatide |
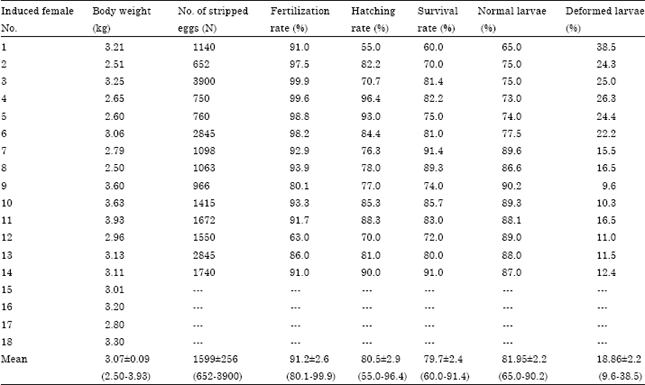 | |
| Data were expressed as Mean±SEM, Values in the brackets represent the range of data (minimum-maximum) | |
Embryonic development: The embryonic developmental stages are shown in Table 2 and Fig. 1. The description of each stage was based on its anatomy.
Fertilized egg: The stripped eggs were spherical in shape, yellowish in colour and semi transparent. Fifteen minutes after fertilization, the ova start to swell and increased its diameter. The size of fertilized eggs ranged from 2.5-3.3 mm with the mean of 2.84±0.09 mm (Fig. 1a). At 2 h after Fertilization (HAF), a small perivitelline space was formed that separated the egg membrane from the rest of the egg (Fig. 1d).
Formation of embryo: The first cleavage in which the blastodisc was divided into 2 blastomeres began at approximately 2 HAF. The next five cleavage cycles occur at regular intervals of approximately 30 min to 1 h (Fig. 1d-g). The egg showed a typical discoidal meroblastic segmentation and formed an equal size of blastomere (Fig. 1d). The second cleavage (4-cell stage) was noticed at 2 h 30 min after fertilization (Fig. 1e) and the 8-cell stage appeared within 3 h 15 min after fertilization. The 16 and 32 cell stages were observed at 4 and 5 h HAF, respectively (Fig. 1f, g). During 32-cell stage, oil droplets appeared but unclear. The blastomeres decreased in size as successive cleavages occurred and the morula stage appeared between 9 and 9 h, 45 min post fertilization (Fig. 1h). At this stage the anterior and posterior ends of embryo have differentiated when the time of the crown of blastomeres started to spread in the form of thin layer over the yolk. The oil droplets gradually began to fuse to form clearer and larger droplets (10-15 droplets). The embryo attained the blastula stage at about 10-14 HAF when the flattening of the cellular material occurred (Fig. 1i). At this stage, it was difficult to count the number of cells and cell division became less synchronous.
| Table 2: | Summary of egg and larval development of the F1 generation of Tor tambroides under normal environmental conditions in captivity (25-28°C) |
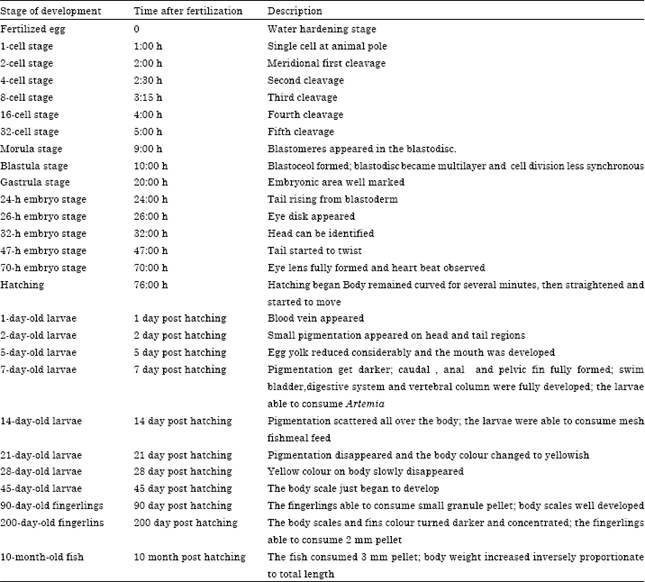 | |
| A total of 50 eggs, 50 larvae, 50 fingerlings and 50 fish were examined | |
Blastodisc became multilayered and the oil droplets decreased in number to 8-10 (Fig. 1i). Gastrula stage was observed between 20-21 HAF when blastoderm expanded as cells proliferated into a sheet and spread toward the vegetal pole (Fig. 1j). The embryonic streak was observed during this stage. The eggs globules became less apparent.
Differentiation of embryo: At 24 HAF, an embryonic body was formed from embryonic streak (Fig. 1k). The eye rudiment and tail could be identified during 26 HAF (Fig. 1l). At 32 HAF, the embryo rose above the yolk and head appeared (Fig. 1m). At this stage, the tail region started to detach from the yolk and ectodermal thickening appeared to form lens of the eye. The tail was detached from the yolk and showed slight movement at 47 HAF (Fig. 1n). At this stage, the body of the embryo covered one-quarter of the yolk and optic vesicles. The anterior part formed a head fold while the posterior part elongated to form a tail fold.
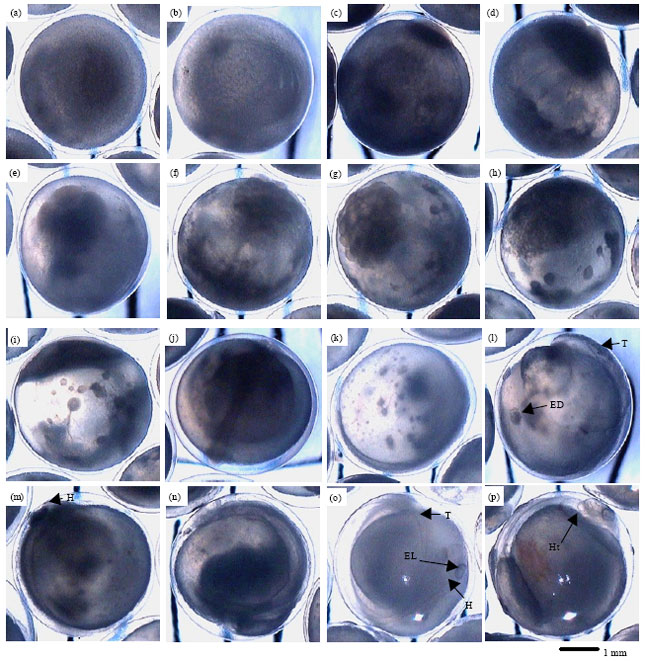 | |
| Fig. 1(a-p): | Cleavage and embryonic development of T. tambroides, (a) 30 min post fertilization egg, (b) Early 1-cell stage 45 min Post Fertilization (PF), (c) One cell stage 1 Ha after Fertilization (HAF), (d) First cleavage (2-cell stage) 2 HAF, (e) Second cleavage (4-cell stage) 2 h 30 min PF, (f) Fourth cleavage (16-cell stage) 4 HAF, (g) Fifth cleavage (32-cell stage) 5 HAF, (h) Multicell morula stage 9 HAF, (i) Blastula stage 10 h PF, (j) Gastrula stage 20 HAF: Embryonic streak visible, (k) 24 h embryo stage; the tail (T) was rising from blastoderm, (l) 26 h embryonic stage: Eye Disk (ED) start to appear, (m) 32 h embryo; head (H) was observed, (n) 47 h embryo; tail started to flip and (o-p) 70 h embryo; eye lens (EL) fully form: Heart (Ht) beat was observed scale bar = 1mm |
The appearance of mouth and fully formed lens in the eyes were clearly observed at 70 HAF (Fig. 1o, p). At this stage, the embryo showed aggressive movements with 40-55 contractions per minute, the heart beat ranged from 130-145 beats min-1 and blood circulation was observed over the yolk. The notochord was prominent and the shape of the brain was noticed. Hatching took place at about 75-85 HAF. Frequent embryonic twitching movements were noticed as the embryos tried to rupture the egg membrane and then eventually the middle part of the embryo had separated from the egg envelop. The hatchling emerged successfully from the egg capsule after the egg membrane had been broken at 74 HAF.
Hatching and larval development: The hatching started approximately 76 HAF, at a water temperature which ranged from 26 and 27.5°C. The newly hatched larvae were transparent and faint yellow in colour. The means of the total length of larvae was 7.44±0.06 mm, with yolk sac of 4.5±0.44 mm long and 2.16±0.13 mm wide (Fig. 2a). At this stage, the yolk sac developed into two forms (primary and secondary yolk sacs) and pale yellowish in colour. The hatchling eyes were unpigmented while the mouth and fins were under developed. At 10-15 h after hatching (HAH), differentiation of fin folds were noticed. A thin layer of membranous fin fold was observed surrounding the caudal region extended up to the yolk sac (Fig. 2a). The larvae swam rapidly and dwelled at the bottom of the incubator tray in an oblique position with head down and belly up. The total length of 1 day old larvae ranged from 10.0-10.7 mm and faint yellow in colour (Fig. 2b). Blood circulation was clearly observed when a functional heart started to pump the blood pass through the yolk into the tail and back. The heart beat ranged from 165-170 beats min-1. Blood corpuscles were reddish orange in colour showing the development of hemoglobin. The yolk sac slowly decreased in mass and only then the larvae could move faster.
The average Total Length (TL) of 2-day-old larvae was 11.5 mm (n = 50); the yolk sac has reduced in size while the mouth started to develop (Fig. 2c). The mouth invaginated but closed and the eye became pigmented. Different parts of alimentary canal such as esophagus, stomach and intestine began to differentiate even though the yolk sac was not yet completely absorbed. Melanophores were scattered on the head and few of them can be seen on the yolk sac, tail and fin fold. The 5-day-old larva was about 12.8 mm in TL (Fig. 2d). A greatly reduced yolk size (5 mm in length and 1.5 mm in width) was observed. More melanophores were observed on the body and eye disk. Some of the larvae started to swim up. An average TL of 7-day-old larvae was 12.8 mm (n = 50) with good development of upper and lower jaws and the mouth was completely opened (Fig. 2e). After yolk diminished, swim bladder was clearly seen. Differentiation of alimentary canal (esophagus, stomach and intestine) was nearly completed and the larva was able to take exogenous live foods, such as Artemia.
The melanophores extended from the head along the trunk and concentrated at the base of caudal fin (Fig. 2e). Caudal, anal and dorsal fins were completely defined (Fig. 2f, g). The average TL of 14-day-old larvae was 15.5 mm (n = 50), with more pigmentation scattered all over the body (Fig. 2i) and at this stage it was able to consume crumble fishmeal base feed. The larvae could swim faster up and down in the rearing tank. An average TL of 21-day-old larvae was 16.8 mm (n = 50); the growth was slow (Fig. 2j) and melanophores started to disappear and the body colour become yellowish. At 28-day-old, the average TL was 17.3 mm (n = 50) with yellowish colour of the body gradually diminished (Fig. 2k), whilst the average TL of 45-day-old larvae was 2.9 cm (n = 50).
 | |
| Fig. 2(a-k): | Early larval development of T. tambroides, (a) First hatched larva at 76 h after fertilization (HAF), 7.5 mm in length, (b) At 24 h after Hatching (HAH) larva at 10.5 mm long, Blood Vein (BV) appeared from heart to tail, (c) At 2 Day After Hatching (DAH) at 11.2 mm with Pigment Cells (PC) appeared along the body, (d) At 5 DAH larvae, 12 mm long, pigmentation increased, Yolk (Y) was completely absorbed, the mouth was not yet opened and was invaginated, (e) At 7 DAH larva, 12.5 mm long, pigmentation gets dark and the larva start to eat Artemia, (f) Caudal Fin (CF) line was observe, (g) Swim Bladder (SB) was fully developed, Anus (A), Anal Fin (AF), Pelvic Fin (PF), Dorsal Fin (DF) and Vertebral Column (VC) were observed; (h) Good development of upper and lower jaws, as well as complete heart (Ht) development, (i) At 14 DAH larva, 15 mm long, pigmentation was scattered all over the body, (j) At 21 DAH larva, pigmentation disappeared and the body changed to yellowish and (k) At 28 DAH larvae, the yellow body colour gradually diminished. Scale bar = 1 mm |
 | |
| Fig. 3(a-d): | Post larvae and fingerling of T. tambroides, (a) 45 Day after Hatching (DAH) larva at 2.7 cm long, (b) 90 DAH fingerling at 6.7 cm long, (c) 200 DAH fingerlings at 12.5 cm long and (d) 10-month-old fingerlings |
At this stage, the larvae started to eat exogenous foods, such as granule pellet containing 50% crude protein (Fig. 3a). The body scale began to develop, however the notochord and viscera were still clearly visible.
Post larval development: The average TL and body weight of 90 DAH post larvae were 7.2 cm and 2.4 g, respectively (n = 50). At this stage the post larvae were fed with small granule pellet containing 50% crude protein (Fig. 3b). Concentrated pigmentation at the base of the caudal fin diminished. The body scales well developed covering all parts of the body. The body colour of fingerlings turns into silver whitish and small dark pigmentation on the scales. The lateral line is formed. The average TL and BW of 200 DAH fingerlings was 12.3 mm and 24 g, respectively (n = 50). At this stage, they were fed with 2 mm pellet containing 32% crude protein (Fig. 3c). The body scales had enlarged. The body scales and fins colour turn darker and more concentrated. The averages of TL and BW of 10-month-old fish were 13.2 cm and 31 g, respectively (n = 50). They were fed with 3 mm pellet containing 32% crude protein (Fig. 3d). At this stage, the fish body weight increased faster than total length (TL).
DISCUSSION
In this study, the breeding performance of T. tambroides females after implanted with Ovaplant then followed by injection with Ovatide was considerably good. The use of Ovatide showed 78% ovulation rate which was much better than the previous study by Ingram et al. (2005) with 55% of ovulation rate by using Ovaprim on the same species. Similarly in other studies, Ovatide was found to be more effective and has successfully induced the ovulation in several species such as Labeo rohita and Cirrhina mrigala as compared to Ovaprim (Dhawan and Kaur, 2004). The use of sustained release delivery of Gonadotropin-releasing Hormone Analogue (GnRHa), such as Ovaplant, advances final oocyte maturation in T. tambroides and has improved the likelihood of ovulation induction (Ingram et al., 2005). Ovaplant has also been successfully applied in a range of fish species to advance final oocyte maturation, ovulation and spermiation (Mylonas and Zohar, 2001). The release of GnRHa may be sustained for up to 6 weeks in slow-release cholesterol-based pellets (Goren et al., 1995). Hence, the effectiveness of Ovaplant and Ovatide were tested in this study.
To the best of knowledge, this is the first successful attempt to use Ovatide as a stimulating agent to induced ovulation in filial 1 (F1) generation of T. tambroides and observation of egg and larval development of filial 2 (F2) generation in a controlled captive condition. In this study, filial 2 (F2) generation of T. tamboides were successfully produced. The present descriptions of embryonic and larval development of T. tambroides are almost similar with other fish, such as European sea bass (Dicentrarchus labrax) (Saka et al., 2001), gilthead sea bream (Sparus aurata) (Kamaci et al., 2005), thin-lipped mullet (Liza ramada) (Mousa, 2010). Duration for the various developmental stages was different might be due to the kind of species and variation of incubation temperatures used. It is a long established fact that egg development and hatching time are both temperature dependent. The time required for fertilized eggs to hatch decreases with the increase of temperature (Mousa, 2010). In this study, the experiments were conducted under the desirable temperature range and mostly at the optimum level for Tor tambroides. The fertilized eggs developed and hatched within 76 h at 25-28°C of water temperature. These conditions were within the range as those in Ingram et al. (2005) whereby under hatchery conditions that the eggs of T. tambroides hatched and developed within 68-82 h at 26-30°C in hatchery conditions.
The fecundity (ranged 260-1200 eggs kg-1), egg diameter before fertilization (2.0-2.8 mm), egg diameter after fertilization (water-hardened eggs 2.5-3.3 mm), time to hatching (from 76 HAF) and the onset of exogenous feeding (7 days) of T. tambroides in this study were comparable with other study on T. tambroides (Ingram et al., 2005) and other Tor species (Nautiyal and Lal, 1985; Joshi, 1988; Nandeesha et al., 1993; Joshi et al., 2002; Ingram et al., 2005). Ingram et al. (2005) also reported that T. tambroides egg diameter before fertilization was 2.20-3.08 mm (mean 2.69 mm) and after 15 min of fertilization, the eggs swelled up to 20%. The fecundity of F1 generation of T. tambroides in this study was lower than the wild caught of T. tambroides in Ingram et al. (2005). The bi-lobed yolk sac observed in T. tambroides embryo is the unique characteristic of the genus. Similar observation was reported in T. kudree and T. douronensis embryos (Kulkarni, 1971; Ingram et al., 2005). The fertilization and hatching rates (80.1-99.9% and 55-96.4%, respectively) observed in T. tambroides were in accordance with reports of similar species by Ingram et al. (2005) and other Tor species by Kulkarni (1971), Joshi (1988) and Ingram et al. (2005). The fertilization and hatching rates in this study were higher than the study conducted by Ingram et al. (2005) which used Ovaprim as an inducing hormone. All stripped eggs before fertilization were firm, translucent and pale yellow in colour. Ingram et al. (2005) stated that the stripped eggs of fish before fertilization were firm and pale yellow to golden orange in colour. The eggs colour can be grouped by Pantone Machine System (PMS) colours and were classed into three categories which are light (PMS120, PMS127, PMS134, PMS135, PMS1345), medium (PMS122, PMS129) and dark (PMS130, PMS143) (Ingram et al., 2005). T. tambroides eggs in the study by Ingram et al. (2005) were ‘dark’ in colour. F1 generation of T. tambroides eggs in this study can be categorized as ‘light’ in colour. The translucent condition of the eggs makes it easier to observe the embryonic development. Ingram et al. (2005) reported the difficulties to observe a clear view of embryonic development within 1 HAF.
At the onset of exogenous feeding at 7 DAH, Tor tambroides larvae can readily accept newly hatched Artemia as their food. However, Ingram et al. (2005) observed that T. tambroides larvae can take exogenous feed (mashed hard-boiled chicken yolk or artificial diet) within 5 DAH. In other studies showed that the highest growth rate of T. putitora larvae was achieved on a natural diet of planktons compared to artificial diets (Rai et al., 1994; Bista et al., 2002). At 14 DAH larvae were given high protein crumble fishmeal in combination with mesh pellet to increase growth rate and to avoid further contamination from Artemia shells. The fish can accept commercial pellet starting from 90 DAH of age. Deformities in hatched larvae (18.86%) in this study is considered low compared to the study conducted by Ingram et al. (2005). The reasons for deformities may be related to inherited genetic disorders, non-heritable birth defects, physical injury, disease, chemicals, nutritional deficiencies in broodfish and environmental conditions, including culture techniques (Bodammer, 1993; Tave and Handwerker, 1994; Leatherland and Woo, 1998; Ingram et al., 2005).
In this study, growth rate for filial 2 generation of T. tambroides was low as also reported by Sehgal (1999) in wild mahseer. The slow growth of T. putitora fingerling, 6-19-23 g in 210 days and 19-23-44 g to 44 g in 6 months was observed when they were reared in low water temperature (Bista et al., 2002). Cement tank condition in closed environment also may attribute to slow growing of the fish compare to the pond-reared fish which exposed to natural foods. Growth rates of pond-reared T. tambroides in the study by Ingram et al. (2005) were consider higher than the present study which reached a mean weight from 142-179 g (maximum 270 g) in 60 weeks.
CONCLUSION
The successful induced breeding of F2 generation of Tor tambroides has been reported for the first time in Sarawak, Malaysia. Ovatide with 0.5 mL kg-1 b.wt. dosage was used to induced ovulation in the female. This study has sequentially described and illustrated the detailed embryonic, larval and post larval development of T. tambroides under rearing conditions. These results have contributed to database of early ontogeny of Tor as one of the important cyprinids in Malaysia and other countries where Tor exist.
ACKNOWLEDGMENTS
Funding of this project was provided by Agri-Biotechnology Institute (ABI). The authors wish to thank Mr. Bill Lu Thian Tack owner of LTT Aquaculture Farm, Asajaya, Sarawak Malaysia for allowing the use of his facilities and the broodstocks. Thanks also go to Raymond Britin, Chai Fook Ngian, Muhammad Zaidi Dini and Chai Nyong Jee for their assistance while sampling. The help provided by staffs at Tarat Indigenous Fisheries Production and Research Center (IFPRC), Serian, Sarawak, is very much appreciated.
REFERENCES
- Azuadi, N.M., S.S. Siraj, S.K. Daud, A. Christianus, S.A. Harmin, S. Sungan and R. Britin, 2011. Enhancing ovulation of Malaysian mahseer (Tor tambroides) in captivity by removal of dopaminergic inhibition. J. Fish. Aquatic Sci., 6: 740-750.
CrossRefDirect Link - Bista, J., B.R. Pradhan, A.K. Rai, R.K. Shrestha and T.B.Gurung, 2002. Nutrition, Feed and Feeding of Gold Mahseer (Tor Putitora) for Domestication and Production in Nepal. In: Cold Water Fisheries in the Trans-Himalayan Countries, Petr, T. and S.B. Swar (Eds.). Food and Agriculture Organization of the United Nations, Rome, pp: 107-117.
- Dhawan, A. and K. Kaur, 2004. Comparative efficacy of ovaprim and ovatide in carp breeding. Indian J. Fish., 51: 227-228.
Direct Link - Goren, A., H. Gustafson and D. Doering, 1995. Field trials demonstrate the efficacy and commercial benefit of a GnRHa implant to control ovulation and spermiation in salmonids. Proceedings of the 5th International Symposium on the Reproductive Physiology of Fish, Volume 95, July 2-8, 1995, Austin, TX., USA., pp: 99-101.
- Ingram, B., S. Sungan, G. Gooley, S.Y. Sim, D. Tinggi and S.S. De Silva, 2005. Induced spawning, larval development and rearing of two indigenous Malaysian mahseer, Tor tambroides and T. douronensis. Aquacult. Res., 36: 983-995.
CrossRefDirect Link - Ingram, B.A., S. Sungan, D. Tinggi, Y.S. Sim and S.S. De Silva, 2007. Breeding performance of Malaysian mahseer, Tor tambroides and T. douronensis broodfish in captivity. Aquacult. Res., 38: 809-818.
CrossRefDirect Link - Joshi, C.B., 1988. Induced breeding of mahseer, Tor putitora (Hamilton). J. Inland Fish. Soc. India, 20: 66-67.
Direct Link - Joshi, P.L., T.B. Gurung, S.R. Basnyat and A.P. Nepal, 2002. Domestication of wild golden mahseer (Tor putitora) and hatchery operation. FAO Fish. Tech. Pap., 431: 173-178.
Direct Link - Mousa, M.A., 2010. Induced spawning and embryonic development of Liza ramada reared in freshwater ponds. Anim. Reprod. Sci., 119: 115-122.
CrossRef - Mylonas, C.C. and Y. Zohar, 2001. Use of GnRHa-delivery systems for the control of reproduction in fish. Rev. Fish Biol. Fish., 10: 463-491.
CrossRef - Rai, A.K., B.C. Shreshtha, B.R. Pradhant, I.S. Gurung and S. Toshida, 1994. Larval Rearing of Mahseer (Tor puttitora) Fed Different Feed Ingredients. In: The Third Asian Fisheries Forum, Chou, L.M., A.D. Munro, T.J. Lam, T.W. Chen and L.K.K. Cheong et al. (Eds.). Asian Fishery Society, Manila, Philippines, pp: 106-107.
- Saka, S., D. Coban and K. Firat, 2004. The study on the technology of producing seabream (Sparus aurata L., 1758) larvae in marine fish hatcheries in Turkey. EU J. Fish. Aquat. Sci., 21: 215-218.
Direct Link - Tave, D. and T.S. Handwerker, 1994. Semi-operculum: A non-heritable birth defect in Tilapia nilotica. J. World Aquacult. Soc., 25: 333-336.
CrossRef - Leatherland, J.F. and P.T.K. Woo, 1998. Fish Diseases and Disorders Vol: 2. Non-infectious Disorders. 2nd Edn., CABI Publishing, Wallingford, London. UK., pp: 395.
Direct Link