
Biswajit Roy
Department of Livestock production and Management,College of Veterinary Sciences and Animal Husbandry, Kuthulia, Rewa, MP-486001, India
B. Brahma
Department of Livestock production and Management,College of Veterinary Sciences and Animal Husbandry, Kuthulia, Rewa, MP-486001, India
S. Ghosh
Department of Livestock production and Management,College of Veterinary Sciences and Animal Husbandry, Kuthulia, Rewa, MP-486001, India
P.K. Pankaj
Department of Livestock production and Management,College of Veterinary Sciences and Animal Husbandry, Kuthulia, Rewa, MP-486001, India
G. Mandal
Department of Livestock production and Management,College of Veterinary Sciences and Animal Husbandry, Kuthulia, Rewa, MP-486001, India
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 1-19







ABSTRACT
Protein is an expensive ingredient of dairy cattle feeds. Overfeeding of protein contributes to environmental pollution and higher feed costs. Although, high dietary protein stimulates milk production, increased protein has been found to be detrimental to reproductive performance of the animal. Efficiency of protein feeding is a function of nitrogen supply to the rumen as well the cow. Consequently, a system to monitor protein feeding must account for rumen as well as post ruminal supply and efficiency. Milk Urea Nitrogen (MUN) has potential to be utilized as a non-invasive input to a system to monitor the crude protein status in a dairy herd on a regular basis. The MUN in a dairy herd provides an opportunity to formulate the dietary protein constituency that optimizes nitrogen utilization for milk production and avoids possible negative effects on herd fertility. Milk urea varied by season, month, parity group, stage of lactation and sample type. Researchers should consider controlling for these variables as potential confounders when exploring the relationship between MU and nutritional management or measures of performance such as production or reproduction.
PDF Abstract XML References Citation
How to cite this article
Biswajit Roy, B. Brahma, S. Ghosh, P.K. Pankaj and G. Mandal, 2011. Evaluation of Milk Urea Concentration as Useful Indicator for Dairy Herd Management: A Review. Asian Journal of Animal and Veterinary Advances, 6: 1-19.
DOI: 10.3923/ajava.2011.1.19
URL: https://scialert.net/abstract/?doi=ajava.2011.1.19
DOI: 10.3923/ajava.2011.1.19
URL: https://scialert.net/abstract/?doi=ajava.2011.1.19
INTRODUCTION
Increased genetic potential for milk production has been associated with a decline in fertility of lactating diary cows. Strategies for meeting the nutritional requirements of high producing cows have necessarily changed in conjunction with genetic gains. Increasing protein concentration of the diet of lactating dairy cows can often increase milk production. However, efficiency of utilization of dietary protein for milk production decreased as more protein was offered. In an effort to sustain milk production during early lactation, dairy producers often increase nutrient density of dairy cows diets to compensate for suboptimal intakes. This situation may lead to protein intakes in excess of recommendations or requirements.
Protein is an expensive ingredient of dairy cattle feeds and thus overfeeding of protein could be costly to producers. Although, high dietary protein stimulates milk production, increased protein has been found to be detrimental to reproductive performance of the animal (Ferguson and Chalupa, 1989; Canfield et al., 1990; Wenninger and Distl, 1994; Guo et al., 2004). Conception and the establishment of pregnancy are an ordered progression of interrelated events involving all of the various tissues of the reproductive tract, follicular development resulting in ovulation, fertilization of the oocyte, embryo transport and development, maternal recognition and implantation. Ammonia, urea or some other toxic products of protein metabolism may intercede at one or more of these steps to impair reproductive efficiency (Jordan et al., 1983).
Another aspects regarding excess protein feeding in dairy cattle is a growing concern for the environment in the last decade. Overfeeding of protein contributes to environmental pollution and higher feed costs (Ferguson et al., 1988; Blanchard et al., 1990; Baker et al., 1995; Burgos et al., 2007). Vandehaar (1998) predicted that the energy loss from feeding an extra 2% of protein (a diet of 19% CP) would amount to 0.36 Mcal day-1. Animal husbandry wastes can contribute to Nitrogen (N) pollution of the environment as ammonia volatized to the air, nitrate leached to ground water and N that runs off to surface water. Improvement of N utilisation efficiency by domestic animals decreases N losses form farms. A majority of manure N applied to fields is subsequently lost to water resources even under the best management condition. Therefore, it is important to reduce manure N output by improving N utilisation by the animal. Furthermore, much fertilizer N applied to crops is also lost to water resources before crop intake. Therefore, the improvement of nutrient utilisation by the animal in an effort to reduce the need for crop production further decreases N losses from animal.
Now-a-days, urea and other Non-Protein Nitrogen (NPN) compound has been used extensively for the feeding of ruminants to fulfill their protein requirements, as the conventional protein sources are expensive. The principle behind this approach is to capitalize the ability of the rumen micro-organisms to convert NPN compounds to protein. Urea entering the rumen is rapidly hydrolysed to ammonia by bacterial urease and the rumen ammonia concentration is therefore liable to rise considerably. For this ammonia to be efficiently incorporated in microbial protein, two conditions must be met. Firstly, the initial ammonia concentration must be below the optimum and secondly, the micro-organism must have a readily available source of energy for protein synthesis. Therefore, urea will not be used efficiently by the ruminants unless Ruminal Degradable Protein (RDP) to satisfy the needs of its rumen micro-organisms. Efficiency of protein feeding is a function of nitrogen supply to the rumen as well the cow. Consequently, a system to monitor protein feeding must account for rumen as well as post ruminal supply and efficiency.
How to monitor protein status of the animals?: Two basic ways to monitor the protein status of the dairy animals are by (1) monitoring intake parameter that is crude protein intake of individual animals and (2) monitoring output parameter that is potentially useful is milk or blood urea concentration.
MUN as an indicator of protein status of dairy animals: We need a tool to monitor protein nutrition status of lactating cows and that would be beneficial to reduce losses and maximize efficiency of nitrogen utilization. Suitable input parameters are not available because protein intake or some related trait is difficult to measure in practice, mainly because of the inaccuracies in predicting feed intake. An output parameter that is potentially useful is milk or blood urea concentration. These are mainly three sources of urea in milk (1) end product of protein and (2) NPN digestion and (3) amino acid catabolism in mammary gland. Results from MUN/BUN measurements can provide valuable information to farmers on the nutritional status and health of their cows. The use of MUN as a fertility marker in dairy cows has received much attention recently (Garcia-Bojalil et al., 1994; Larson et al., 1997).
What is milk urea nitrogen (MUN)?: Milk protein consists of two major protein fractions-true protein and NPN. True protein accounts for 95% of the nitrogen in milk and it consists of 80% casein and 20% whey. The NPN makes up the remaining 5% of the total nitrogen in milk. The NPN consists of approximately 30-35% milk urea, 25% creatinine/uric acid, 15% amino acids and 10-30% ammonia.
Physiological basis: Rumen ammonia nitrogen in excess of rumen requirement usually is the single largest contributor of urea in the blood; however, catabolism of body protein and deamination of excess dietary protein can contribute to the pool of urea nitrogen in the blood. Rumen ammonia concentration in excess of that utilized by or captured by rumen microbes is absorbed across the rumen wall into the portal blood. Rumen ammonia concentration and rumen pH are the two major factors which effect level of ammonia exchange with portal blood. Normal ruminal pH is 6.0 to 6.8 where ammonia exists mostly as ammonium ion (NH4+) and exchange with the blood is uncharged ion (NH3) through passive diffusion. Therefore, feed or feeding factors elevate rumen ammonia concentration (i.e., high dietary soluble protein) or pH (i.e., high roughage diets), increase transfer of ammonia to the blood.
Excess amino acids and peptides are deaminated in the liver and N is converted to urea. Ammonia, because it is toxic to the animal, is rapidly converted to urea in the liver (Swenson and Reece, 1993). Normal blood ammonia (0.1-0.15 mg %) is low relative to rumen levels (10-50 mg %) due to the rapid detoxification of blood ammonia to urea in the liver. Estimates of the energy costs to synthesize and eliminate excess urea have been calculated to around 12 Kcal of DE per gram of urea synthesized (Van Soest, 1994). The urea enters the circulatory system through the hepatic sinuses, which drain into the hepatic vein (Guyton, 1982) and become part of the pool of blood urea N. The urea is filtered from the blood by the kidney and is excreted from the body in urine. Blood enters the kidneys through the renal artery (Guyton, 1982) and is filtered through the nephrons. This process concentrates the urea for excretion in the urine. Because of counter current flow and differences in membrane permeability in ascending and descending loops of Henle, a concentration gradient for the diffusion of urea into urine is created to remove urea from blood (Swenson and Reece, 1993). Blood flow through the kidney is constant within an animal, which ensures a constant urea filtration rate (milliliters of blood filtered per minute), regardless of urine volume. With a low volume of urine, urea concentration in the urine would be higher than with a higher volume of urine, but a similar amount of blood would be cleared of urea (Swenson and Reece, 1993). In addition, with high concentrations of urea in blood, more urea would be removed per minute compared with a low concentration of urea in the blood, but the total amount of blood cleared would remain similar. Therefore, urea excretion is proportional to blood urea concentration. Urea, because it is a small neutral molecule, readily diffuses across cellular membranes. As milk is secreted in the mammary gland, urea diffuses into and out of the mammary gland, equilibrating with urea in the blood. Because of this process, MUN equilibrates with and is proportional to blood urea N (Roseler et al., 1993). This process allows MUN to be an excellent predictor of blood urea N and Urinary N (Kohn et al., 1997; Ciszuk and Gebregziabher, 1994).
Advantages of milk urea estimation over blood urea: While MUN is only indirectly related to ruminal N utilization, depending on the urea clearance rate of the kidneys (Kauffman and St-Pierre, 2001; Kohn et al., 2002), milk is a much more practical fluid to obtain for wide-scale urea testing than urine or blood. As a result, MUN testing can be incorporated as one of the monthly milk tests. Because urea equilibrates within bodily fluids, MUN should be similar to PUN as an indicator of urea nitrogen status in dairy cows and is more conveniently monitored (Oltner and Wiktorsson, 1983; Roy et al., 2005).
Milk Urea Nitrogen (MUN):
| • | Corresponds to 2.5 to 3.0% of total milk N (De Peters and Cant, 1992) |
| • | Is highly correlated (0.88 to 0.98) with Blood Urea Nitrogen (BUN) and is closely related to dietary CP (Butler et al., 1996; Dhali, 2001; Roy et al., 2005) |
| • | Does not require invasive sampling or restraint of the cow |
| • | Non-invasive, economical, rapid process |
| • | Easily sampled and easy to perform |
| • | When milk samples are taken as a part of regular testing, sampling involves no extra labor and it is cheaper than sampling and analyzing blood |
When a group of cows has a mean MUN concentration that is higher than the target MUN values at a given level of production, excess protein for the given level of production was probably consumed. A reformulation of the diet at that production level with a lower protein concentration could reduce feed costs. However, before diet reformulation occurs, the specific cause of high MUN should be identified. High MUN can result from a number of nutritional factors, including but not limited to excess protein, inadequate energy, or excess RDP that decreases production and efficiency of N utilization. Close examination of the current diet helps to elucidate the cause of high MUN and leads to appropriate dietary changes to reduce MUN.
Individual MUN vs. Bulk tank MUN estimation: It has been suggested that the mean MUN concentration from a group of cows should fall within a specific range, with values outside that range reflecting nutritional problems (Broderick and Clayton, 1997; Jonker et al., 1998). An alternative to determining the mean MUN level for all the cows in a herd is to test the bulk tank MUN (BTMUN). The cost and labor advantages of testing one BTMUN sample versus many individual cows seems attractive. However, it is still unclear just how reliable BTMUN is as a guide to the average urea concentration of a herd.
Theoretically, BTMUN should be equal to a weighted (by milk production) herd average of individual MUNs (WHMUN) if individual milk weights, individual cow MUNs and BTMUN are all simultaneously measured without error. However, potential differences between BTMUN and WHMUN can arise from (1) imprecision in individual milk weights, (2) sampling variance and (3) analytical variance and 4) time effects (BTMUN and individual cow MUNs are not necessarily from the same milkings or same number of milkings). Also, because BTMUN is the combination of many cows’ contributions, with varying MUN values, milk weights and group management; it is difficult to determine if a group of cows has a MUN problem if another group of cows is obscuring the MUN problem.
There are some data on the mean, standard deviation and range of BTMUN in different countries: Sweden (Gustafsson and Carlsson, 1993), Norway (Ropstad and Refsdal, 1987) and Chile (Wittwer et al., 1999) were 11.2±2.1 (4.76 to 17.08) mg dL-1, 13.72±3.36 (4.2 to 32.48) mg dL-1 and 13.16±3.64 (3.92 to 28.0) mg dL-1, respectively. No data are available from Indian context. Two studies have looked at the correlation between BTMUN and weighted herd average blood urea nitrogen and have reported Pearson correlation values of r = 0.77 (Refsdal, 1983) and r = 0.95 (Wittwer et al., 1999). Nelson (1996) stated that a high normal value of BTMUN was difficult to interpret. Bulk tank samples have been observed to differ from the mean of all cows by as much as 3.0 to 4.5 mg dL-1 (Nelson, 1996). Therefore, it is necessary to investigate more thoroughly whether a BTMUN can be a reliable measure of the average milk urea concentration of a herd and whether this relationship is affected by other factors such as herd size, type of sampling that is used on the farm and season of the year.
The seasonal variation in BTMUN also needs investigation. There are few studies that have looked at seasonal variation of BTMUN. It is likely to vary seasonally in the same manner that individual cow MUN values vary seasonally (Arunvipas et al., 2003). Feed changes associated with season, such as the use of corn silages, fresh silage and pasture, are likely to influence MUN values. However, there needs to be documentation of this seasonal variation. Also, the relationship between BTMUN and WHMUN could be affected by season. For example, different groups of cattle may be managed differently on pasture, with high producing cattle given better quality pasture, or with access to pasture given at different times to different groups. Therefore, BTMUN could be different from WHMUN during specific times of the year. The BTMUN is inherently a whole herd assessment of MUN status without the ability to make conclusions regarding subgroups of cattle within a herd (Arunvipas et al., 2004). There was no relationship either between bulk milk urea concentration and fertility, or between changes in bulk milk urea concentration and fertility (Cottrill et al., 2002). However, bulk tank milk urea concentrations have been used to predict protein supply (Refsdal et al., 1985; Ropstad et al., 1989) and fertility differences among herds (Ropstad and Refsdal, 1987).
Low MUN: Conditions can exist where MUN levels may actually be low indicating a protein deficiency in the diet and potentially lost production. Low MUN levels suggest the cows’ diet does not contain adequate available protein. When MUN levels are extremely low, production may be limited because of a protein deficient diet. Suspicious feeds should be analyzed for acid detergent insoluble nitrogen or bound protein by an animal nutrition laboratory.
MUN in relation to reproduction: Fertility is a major contributor to profitability of the dairy herd (Martin, 1992) and is a trait with a very low heritability value (Hoeschele, 1991). Nutritional management plays one of the most important roles in achieving reproductive goals (Ferguson and Chalupa, 1989). The benefits of feeding excess protein to dairy cattle to maintain peak milk production should be compared with potential negative effects on fertility before such a program is implemented. High protein may affect the ovarian function in several way and in turn the fertility of the animal (Fig. 1). Several studies reported the negative effects of BUN or MUN on reproductive performance in dairy cows and suggested that overfeeding CP caused reproductive stress (Ferguson et al., 1993; Rajala-Schultz et al., 2001). However, others did not find such negative effects of high MUN on fertility of cows (Carroll et al., 1988; Godden et al., 2001).
Jorritsma et al. (2003) have reviewed the influences of urea and ammonia on reproduction during early lactation. Cows within herds with high MUN were associated with reduced probability conception at first service, but not at subsequent services (Ferguson et al., 1993). A negative effect of MUN on conception rate at first service among cows within herds, but no such effects were found at second and third service. In among-herd analyses, MUN had minimal effect on conception rate, of but was associated with greater days open among high-producing herds.
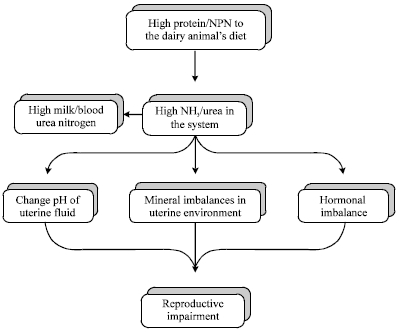 | |
| Fig. 1: | Flow diagram of how milk urea/blood urea relates to reproduction in dairy animals? |
These results agree with the hypothesis that urea affects cleavage and blastocyst formation but not necessarily early oocyte development (Jorritsma et al., 2003). Urea nitrogen concentrations greater than 19 mg dL-1 in plasma and milk were associated with decreased pregnancy rate in dairy cattle (Butler et al., 1995). Similarly, Ferguson et al. (1988, 1993) reported a decrease in conception rate when serum urea nitrogen concentrations were greater than 20 mg dL-1. Rajala-Schultz et al. (2001) found that cows with MUN concentrations below 10 mg dL-1 were nearly two and a half times more likely to be confirmed pregnant than cows with MUN concentrations above 15.4 mg dL-1.
Three general mechanisms have been proposed to describe how excess dietary protein may negatively influence fertility:
| • | Nitrogen by-products may alter the uterine pH and mineral balance |
| • | Nitrogen by-products or efficiency of energy utilization may alter gonadotropin and (or) progesterone secretion |
| • | Toxic by-products of nitrogen metabolism from the rumen (ammonia) and liver (urea) may impair sperm, ova, or early embryo survival |
| • | These effects may occur singularly, simultaneously, or synergistically |
Change of uterine pH and mineral balance: Feeding excess crude protein to dairy cows, regardless of protein source and degradability, alters the uterine environment. How this effect is mediated is not completely known at present. The changes in uterine pH observed in cows fed high levels of crude protein may be linked to reductions and fertility, as reported in other studies. Composition of the uterine luminal fluid in cows fed high protein diets has been examined by several workers to elucidate the mechanisms responsible for reduced conception rates.
Uterine luminal pH is controlled by Carbonic Anhydrase (CA), which catalyzes the reversible reaction: H2O+CO2↔H2CO3↔H++¯HCO3. This enzyme is present in many secretory epithelia including the reproductive tract (Rodriguez-Martinez et al., 1991) and functions in the selective transport of hydrogen and bicarbonate ions (Swenson, 1991). Depending on the permeability of apical vs. basolateral plasmalemmae, epithelial cells can export H+ or HCO3 apically or basally in exchange for sodium, potassium and chloride ions to affect cellular ion content and to modify the pH of the luminal fluid (Rodriguez-Martinez et al., 1991). Unique isozymes of CA are induced by progesterone in the human or rabbit uterus (Hodgen and Falk, 1971; Falk and Hodgen, 1972) and, therefore, abundance varies by stage of cycle. In one study (Rhoads et al., 2004), uterine endometrial biopsies were collected from lactating cows and CA increased between estrus and d 9 of the estrous cycle (4.6±0.4 and 6.1±0.5 units mg-1 of protein, respectively). The apparent steroid-dependency of CA is particularly intriguing because the observed changes in uterine pH in association with PUN occurred only during the luteal phase (Elrod and Butler, 1993; Elrod et al., 1993). Conceivably, high urea could alter uterine pH by changing CA activity during the luteal phase, but not at estrus.
Effects of protein diet on potassium, phosphorus and pH were only observed during the luteal phase. Although, consequences of altered mineral concentrations on embryonic function are not known, a reduction in pH from 7.2 to between 6.9 and 7.1 has been reported to compromise embryonic development in mice (Edwards et al., 1998). In sheep, uterine pH was significantly affected when PUN exceeded 19 mg dL-1. When fed excess rumen degradable protein, PUN concentrations greater than 18 mg dL-1 were detrimental to the early development and survival of embryos (Bishonga et al., 1994). Composition of the uterine luminal fluid in cows fed high protein diets has been examined to elucidate the mechanisms responsible for reduced conception rates. Jordan et al. (1983) found that cows fed a high CP diet had decreased concentrations of P, Mg and Kin uterine fluid during the luteal phase, but uterine P, Mg and K were similar at estrous. However, there was no relationship between elevated systemic concentrations of ammonia and urea and the biochemical composition of oviduct fluid in cattle, with the exception of calcium (Kenny et al., 2001). Elrod and Butler (1993) and Elrod et al. (1993) fed diets differing in RDP and RUP content to dairy cows and heifers, resulting in elevated PUN. Uterine luminal pH was measured on the day of estrus and 7 d after estrus. Uterine pH of control and high PUN cattle was similar at estrus. On d 7 after estrus, however, high PUN concentrations were associated with low uterine pH. Uterine pH on d 7 after estrus is of interest because if the oocyte had been fertilized, the embryo would be within the uterine lumen. Because placentation has not yet occurred, the d 7 embryo is dependent on uterine secretions for survival. Changes in the uterine environment during this period could compromise early development or viability of the embryo. They also reported that for increase in 5 units of PUN (mg dL-1), uterine pH would decrease by 0.5 units. A decrease in uterine luminal pH occurred in response to urea infusion (Rhoads et al., 2004). Lowering of pH in the endometrium is highly sensitive to embryo and may lead to death.
Hormonal imbalance: Progesterone is important for follicular development, passage of the fertilized embryo through the oviduct to the uterus and maintaining pregnancy. The amount and source of protein may influence progesterone (Swanson, 1989). It is possible that decreases in serum progesterone occur at concentrations of dietary crude protein that exceed rumen requirement for rumen degradable protein. However, source of protein, energy status and age of the animal should be considered. Nonetheless, the effect of protein consumption on progesterone concentration needs more study and factors such as total energy intake and protein source should be examined.
Plasma progesterone concentrations during mid-diestrus were reported to be approximately 30 percent lower in cows with high PUN due to feeding high protein diets (Sonderman and Larson, 1989). Whereas, plasma progesterone concentrations did not differ during the 5-d period following AI (early diestrus) when cows were categorized into high and low PUN groups (Butler et al., 1996). Larson et al. (1997) found that non-pregnant cows with low progesterone post breeding were often associated with high MUN.
Habib et al. (1980) observed that presence of Zn or protein decreased the binding capacity of P4 (progesterone) with its receptor in the human endometrium cell culture because Zn-P4-protein complex may compete for the same site with P4. Jordan et al.(1983) observed that high protein in the diet of lactating cows decreased the uterine secretion of K, P and Mg in the postpartum estrous cycle (0-1, 5 and 15th d) but had higher urea (6.4 vs. 17.2 mg dL-1) and Zn concentration. They also reported that high protein decreased the progesterone (P4) concentration on the same period of the cycle. In contrast, Blauwiekel and Kinacid (1986) reported that dietary protein had no effect on pituitary or ovarian hormones in cows. Figueroa et al. (1992) observed high or low UDP (40 and 35%) did not affect the plasma P4 or cervical mucous urea concentration in cows. Vasantrao (1993) reported that in goats high RDP (70%) group had lower P4 than lower RDP group or 20% less CP with high UDP. Sonderman and Larson (1989) observed that high CP had lower circulating peak P4 (3.47 and 4.91 ng mL-1). Presence of urea may diminish the effects of progesterone and large amounts of prostaglandin (PGF2á) may secrete from endometrium, which ultimately interferes with embryo development and viability (Gilbert et al., 1996). Other hypotheses are, to detoxify the high NH3 to urea, energy is diverted and that may aggravate an energy shortage postpartum, such that, metabolic attention is diverted from ovarian activity. A reduction in the capacity of the liver to synthesize glucose from propionate in presence of excess NH3 has been observed (Overton et al., 1998). Plasma glucose was increased and insulin was depressed in cows fed high amounts of RDP (Garcia-Bojalil et al., 1998) and insulin was increased with high UDP in the diet of lactating cows (Chaturvedi and Walli, 2000). Decreased production or utilization of glucose in case of high RDP can result in greater use of body reserve for milk production. This impairment of glucose metabolism may affect the ovarian function (Staples et al., 1998).
Observations by researchers indicated that gonadotropin secretion was not directly affected by diets with high crude protein, but ovaries and thus steroid secretion were affected. Change in steroid secretion then affected the brain and gonadotropin secretion (Canfield et al., 1990). Thus, alteration of the uterine environment as a consequence of high protein may explain the fertility problems.
A system of endometrial cell culture has been developed to study the effects of urea on secretory function (Gilbert et al., 1996). In this three-dimensional system, the endometrial cells become polarized and establish a pH gradient between apical and basal compartments as soon as the cells reach confluence. The pH gradient is sensitive to both estradiol and progesterone. The presence of urea significantly (p<0.05) diminished the effects of progesterone in maintaining a pH differential between apical and basal compartments. Large amounts of PGF2α and PGE2 are secreted by the endometrial cultures. Treatment of the cultures with both estradiol and progesterone suppressed PGF production, but the presence of urea significantly (p<0.01) increased the secretion of PGF2α and PGE2. The importance of further studies on the effects of urea on the production of endometrial prostaglandin stems from the evidence that PGF2α interferes with embryo development (Mauer and Beier, 1976) and viability (Schrick et al., 1993).
Gamet and embryo survivality: Embryonic mortality is a limiting factor to the success of reproduction in cattle. Most pregnancy wastage occurs early during the pregnancy and the causes are usually unknown. Oviductal fluid provides nutritional environment for the early embryo, the metabolite, facilitates gamete transport and maturation, fertilization and therefore ionic composition of oviductal fluid is clearly important. Early embryonic development requires appropriate oviductal and uterine environments. Variations in the uterine environment caused by high PUN concentrations may therefore create a hostile or suboptimal environment for early embryo development. High PUN concentrations in lactating dairy cows decrease embryo viability through effects exerted on the oocyte or embryo before recovery from the uterus 7 days after insemination (Rhoads et al., 2006).
Threshold systemic urea concentrations of 5.7 and 6.8 mmol L-1, above which a 30 and 20% reduction in pregnancy rate for Holstein heifers and cows, respectively, was observed (Elrod and Butler, 1993; Butler et al., 1996). Urea concentrations are elevated in uterine secretions of cows fed large amounts of CP (Jordan et al., 1983) and in oviductal-uterine flushings of ewes fed supplemental urea (McEvoy et al., 1997).
Excess rumen degradable protein has been reported to be deleterious to embryonic development in lactating cows (Blanchard et al., 1990), but not in non-lactating cows (Garcia-Bojalil et al., 1994). Uterine secretions altered in high-producing cows fed high crude protein diets and resulting in high PUN (Jordan et al., 1983). However, no relationship was found between elevated systemic concentrations of ammonia and urea and the biochemical composition of oviduct fluid in cattle, with the exception of calcium (Kenny et al., 2001). It is likely that feeding excess amounts of CP also elevates urea concentrations in the follicle, the site of oocyte maturation (Ocon and Hansen, 2003). Anderson and Barton (1987) suggested that elevation of tissue ammonia associated with high intakes of crude protein may delay clearance of uterine contaminants by reducing immune system function. In herds which experience reproductive problems with early postpartum cows, feeding a high crude protein diet containing high rumen degradable protein, may decrease the fertility, unless the herd health program provides for early identification and treatment of problem cows.
Exposure of oocytes to physiologically relevant concentrations of urea during the process of maturation interfered with ability of the embryos formed after fertilization to develop to the blastocyst stage (Ocon and Hansen, 2003). Exposure of oocytes to 6 mM of urea during maturation in vitro impaired meiosis and fertilization rate (De Wit et al., 2001). Feeding a high CP diet increased growth of the second-wave dominant follicle, decreased capacity of oocytes from small (1 to 4 mm) and medium (>4 to 8 mm) follicles to cleave when fertilized in culture and decreased ability of cleaved embryos formed from oocytes obtained from medium-sized follicles to develop in culture to the blastocyst stage (Sinclair et al., 2000). Exposure of the embryo to urea after fertilization had no effects on development, indicating that the embryo itself was resistant to direct effects of urea (Ocon and Hansen, 2003). Embryo survival rate is not affected by either CP or fermentable carbohydrate intake, irrespective of whether animals are fed a silage diet supplemented with feed grade urea or are grazing pasture with a high crude protein content (Kenny et al., 2000). In contrast to its resistance to urea, the embryo was very susceptible to disruption by reduction in pH, suggesting that the reduction in uterine fluid pH in the luteal phase of cows fed a high-protein diet (Elrod et al., 1993; Elrod and Butler, 1993) is inimical to embryonic survival. Taken together, the results implicate changes in urea concentrations and uterine pH as a cause of infertility associated with feeding diets containing excess CP.
However, Gath et al. (1999), in an embryo-transfer study, reported that despite generating systemic urea concentrations of more than 7 mmol L-1, neither fertilization rate nor subsequent embryo survival rate in recipient heifers was compromised. Similarly, two recent studies at this laboratory (Kenny et al., 2001), despite generating systemic urea concentrations of up to 25 mmol L-1, failed to establish any detrimental effect on embryo survival rate. Consistent with these findings, the results of the present study show that even with systemic and oviductal concentrations of urea peaking at more than 14 mmol L-1, with the exception of calcium, no relationship is observed between urea and any oviductal or systemic metabolite or ion measured. Similarly, Berardinelli et al. (1997) reported that feeding excess RDP to ewes generated significant increases in systemic concentrations of urea but did not affect either oviductal or uterine protein secretion in the early postovulatory period. The high oviductal concentrations of ammonia recorded in the control animals, suggest that bovine gametes, as part of their normal environment, are accustomed to relatively high concentrations of ammonia. Leese et al. (1979) reported physiological follicular ammonia concentrations up to 366 mmol L-1 during the in vivo development of bovine oocytes.
Non-nutritional factors affect the MUN/BUN level: Majority of the aforementioned studies were performed on individual animals under experimental conditions using chemical tests to measure MU. The association between MU and both nutritional management and performance should be determined under field conditions using commercial testing procedures (Sackett et al., 1991). To investigate these associations, a measure of the non-nutritional factors affecting MU is needed.
Association between milk urea and season, sampling and cow factors: High MUN may be caused by many factors. Excessive protein intake is a common nutritional factor (Jonker et al., 1998). Blood urea nitrogen or plasma urea nitrogen, which is the origin of MUN, may also be affected by diseases or medicines from treatments (Vestweber et al., 1989). Any disease or body condition that reduces glomerular filtration such as dehydration, heart disease and renal disease or any condition that increases protein catabolism can result in increased blood urea nitrogen level (Fraser, 1991).
Any number of factors including health or energy balance can affect MUN among individual cows within a herd (Collard et al., 2000; Stockham and Scott, 2002).
Milk production effect: During early lactation, dietary energy intake does not meet energy requirements for increasing milk production. As a result, body fat is mobilized. Reports on the association between MU and milk yield vary between positive (Carlsson et al., 1995; Kung and Huber, 1983; Oltner et al., 1985), no association (Baker et al., 1995; Carroll et al., 1988) and negative (Ismail et al., 1996). The positive association between MU and milk yield may be attributed to increased milk production which resulted from increased levels of dietary protein fed (Chalupa, 1984; Oldham, 1984). Supplemental protein may increase milk yield by providing more AA for milk protein synthesis, by increasing the available energy through deamination of AA, or by altering the efficiency of utilization of absorbed nutrients (Chalupa, 1984).
There was a positive association between MU concentration and milk yield (Rajala-Schultz and Saville, 2003) for high-producing herds. Other studies examining the relationship between milk yield and MU level found either no significant correlation between these parameters (Gustafsson and Carlsson, 1993; Butler et al., 1996; Godden et al., 2001) or a negative link (Broderick and Clayton, 1997; Roy et al., 2003).
Milk component production: The negative nonlinear association between MU and both milk fat and total protein percentages, while statistically significant, was numerically very small, posing the question of their biological and economic significance. Other studies have reported no association between MU and either milk fat or true protein percentages (Jaquette et al., 1986; Klusmeyer et al., 1990). As this study did not measure true protein levels, direct comparisons cannot be made with former studies.
Milk fat percentage: There was a consistent positive association between MU and milk fat content (Godden et al., 2001; Rajala-Schultz and Saville, 2003) for high-producing herds. A possible explanation for this association could be that high amounts of NDFf may increase milk fat content and at the same time raise MU levels because of the high degradability of its protein. Other studies have reported no association between MU and either milk fat percentages (Jaquette et al., 1986; Klusmeyer et al., 1990).
Milk total protein percentage: There was a negative association between MU and milk total protein content (Ferguson et al., 1997). The inverse relationship between MU and milk total protein pinpoints the alternative pathways that N can follow: incorporation into milk protein or excretion as urea. However, other studies (Klusmeyer et al., 1990; Godden et al., 2001; Rajala-Schultz and Saville, 2003; Sharma et al., 2009) found no significant relationship between milk total protein and MU.
Parity: The MU was lower in first-lactation heifers (Oltner et al., 1985; Butler et al., 1995; Carlsson et al., 1995; Rajala-Schultz and Saville, 2003); however other studies reported only minor differences, with no significant association between parity group and MU (Canfield et al., 1990).
Stage of lactation: The results of this study are consistent with previous studies that reported MU varied by stage of lactation (Bruckental et al., 1980; Carlsson et al., 1995). Carlsson et al. (1995) reported that MU was lowest immediately after calving, increased to a maximum between 3 and 6 month of lactation and slowly declined in later lactation. There was significant interaction between parity and DIM. The rate of decline in MU from mid to late lactation was greater in animals in parity two or greater. Changes in ration nutrient composition or feeding programs that occur among parity groups and different stage of lactation could contribute to the variation observed in MU. There may also be physiological or behavioral differences that could affect MU.
Lower MU in first-parity animals may be attributable to lean tissue growth and a correspondingly higher efficiency of AA utilization. As a result, deamination of AA and subsequent urea formation in the liver may be reduced (Oltner et al., 1985). Also, differences in DMI, rumen microbial adaptation and rumen absorptive capacity could contribute to differences in MU at different stages of lactation. However, Schepers and Meijer (1998) found no association between parity or stage of lactation and MU when feeding trials were controlled for nutritional factors. This suggests that non-nutritional factors are of minor importance in explaining the association between MU and either parity or stage of lactation.
Season: High MU observed during the summer months has been reported by Carlsson et al. (1995) and Ferguson et al. (1997). Total protein and true protein (mostly casein) in milk are lower during the summer months, while NPN, which includes urea, increases (Carlsson et al., 1995; Ferguson et al., 1997; Van Soest, 1994). Moller et al. (1993) attributed variation in MU to seasonal changes in pasture protein and energy components. Australian spring pasture contained 20 to 30% CP and 5 to 20% soluble carbohydrate, thus, creating a high protein:energy ratio, which could result in elevated MU. Studies are lacking that describe the effects of non-nutritional factors associated with season (climate, water intake, DMI, or stage of lactation) on MU. Because of the absence of data on nutrient balance at the cow-level in this study, the variation in MU across parity groups, stage of lactation, or seasons could not be explained by nutritional management or non-nutritional factors.
Diurnal variation: Differences between a.m. and p.m. MU concentrations have been reported (Broderick and Clayton, 1997). Others found that a.m./p.m. MU differences may be influenced by differences in feeding to milking intervals between a.m. and p.m. milkings (Godden, 1998). This supports a previous finding that MU was highest when the cow had eaten within 5 or 6 h before sample collection and began to fall as the feeding-to-sampling interval increased (Gustafsson and Palmquist, 1993). The short feeding-to-milking interval (0 to 6 h) typical of the p.m. sampling period in dairy herds and the longer feeding-to-milking interval of the a.m. sampling period could explain the lower MU in the a.m. milk samples in many herds (Godden, 1998).
Because urea equilibrates across the mammary epithelium (Gustafsson and Palmquist, 1993), little variation was found in MUN concentrations in different milk fractions collected during milking (Butler et al., 1996; Roy et al., 2004; Carlsson and Bergstrom, 1994).
Thus, it should make little difference whether a milk sample intended for MUN analysis comes from a composite milk sample or from a quarter strip sample (Roy et al., 2003), before or after milking (Gustafsson and Palmquist, 1993; Carlsson and Bergstrom, 1994).
Gustafsson and Palmquist (1993) and Elrod and Butler (1993) reported that PUN concentrations fluctuate throughout the day. Generally, the minimum PUN concentration is before feeding and the maximum is approximately 4 to 6 h after feeding. Because there is a lag of approximately 1 to 2 h between PUN and MUN peaks (Gustafsson and Palmquist, 1993), sampling time relative to feeding can be important in the interpretation of PUN and MUN measurements, especially when cows are fed dietary forages and concentrates separately rather than a total mixed ration.
Somatic Cell Count (SCC): A strong negative relationship was found between MU and SCC. The SCC followed MU concentrations (examined monthly) in an inverse order (Godden et al., 2001; Rajala-Schultz and Saville, 2003; Hojman et al., 2004). Milk urea is related to protein and NPN supply and their utilization rate in the rumen; SCC reflects the degree of irritation in the udder.
One study of Quebec herds reported a positive association between cow-level SCC and true protein content. They reported a small but significant positive association between SCC and milk NPN levels (which includes urea) (Ng-Kwai-Hang et al., 1985). De Peters and Ferguson (1992), in a review reported that milk from mastitic glands was lower in casein and higher in non-casein protein. They suggested that casein breakdown products contributed to the whey protein fraction of mastitic milk. Although, a negative relationship between cow-level SCC and MU was observed, MU data should be interpreted at the group-level and not the individual cow level (Broderick and Clayton, 1997; Cannas et al., 1998; Kolver and MacMillan, 1993; Oltner et al., 1985; Schepers and Meijer, 1998). Therefore, unless a large proportion of cows in the group or herd have very high SCC, then the herd average MU should not be greatly affected by SCC. In this study, herd average LS was not associated with herd average MU.
Godden et al. (2001) and Morjorie and Lee (1996) reported that MUN values were associated with log SCC values and MU values were lowest for samples with largest SCC and log SCC. Licata (1985) reported that milk from quarters positive to the California mastitis test was 0.45 mM lower in MU content than that from healthy quarters. Roy et al. (2001) indicated that as the intensity of infection increased from mild to moderate, the MU concentrations decreased. This may be due to the intramammary infection that alters the permeability of the udder tissues.
Preservation of milk sample for urea estimation: In azide preserved cow milk or cow milk storage at 4°C, the milk urea was unchanged for one week, but urea gradually decreased in milk at room temperature as reported by Miettinen and Juvonen (1990). Carlsson and Bergstrom (1994) reported that without preservative the urea concentrations in cow milk were not changed significantly after storage during 10 days at 4°C. But when a preservative (bronopol) was added, the urea concentrations remain unchanged till 17 days. Deep freezing did not influence the urea concentration. Urea concentrations in deproteinized cow milk (Dhali, 2001) and buffalo milk (Roy et al., 2004) were not changed upto 30 days when preserved at 4°C. Being able to use preservative is beneficial in situations, such as field trials, when there may be a long period between the time of milk sampling and MUN analysis. Field trials or on-farm MUN analysis would be benefited by an easy and reliable test system.
CONCLUSIONS
Maintaining and monitoring MUN in a dairy herd provides an opportunity to formulate the dietary protein constituency that optimizes nitrogen utilization for milk production and avoids possible negative effects on herd fertility. The MUN is economical, easy to monitor on a monthly basis and could be performed on all animals in a herd.. Maintaining and monitoring MUN in a dairy herd provides an opportunity to formulate the dietary protein constituency that optimizes nitrogen utilization for milk production and avoids possible negative effects on herd fertility. Milk urea varied by season, month, parity group, stage of lactation and sample type. Researchers should consider controlling for these variables as potential confounders when exploring the relationship between MU and nutritional management or measures of performance such as production or reproduction. Because of the apparent effect of a.m. and p.m. sampling on MU concentration, producers on an alternating a.m./p.m. test schedule should test routinely to establish a herd pattern for MU and/or submit the same sampling time consistently.
REFERENCES
- Arunvipas, P., I.R. Dohoo, J.A. van Leeuwen and G.P. Keefe, 2003. The effect of non-nutritional factors on milk urea nitrogen levels in dairy cows in Prince Edward Island. Prev. Vet. Med., 59: 83-93.
CrossRef - Arunvipas, P., J.A. VanLeeuwen, I.R. Dohoo and G.P. Keefe, 2004. Bulk tank milk urea nitrogen: Seasonal patterns and relationship to individual cow milk urea nitrogen values. Can. J. Vet. Res., 68: 169-174.
PubMedDirect Link - Baker, L.D., J.D. Ferguson and W. Chalupa, 1995. Responses in urea and true protein of milk to different protein feeding schemes for dairy cows. J. Dairy Sci., 78: 2424-2434.
CrossRefPubMedDirect Link - Blanchard, T., J. Ferguson, L. Love, T. Takeda, B. Henderson, J. Hasler and W. Chalupa, 1990. Effect of dietary crude-protein type on fertilization and embryo quality in dairy cattle. Am. J. Vet. Res., 51: 905-908.
PubMedDirect Link - Blauwiekel, R. and R.L. Kincaid, 1986. Effect of crude protein and solubility on performance and blood constituents of dairy cows. J. Dairy Sci., 69: 2091-2098.
Direct Link - Broderick, G.A. and M.K. Clayton, 1997. A statistical evaluation of animal and nutritional factors influencing concentrations of milk urea nitrogen. J. Dairy Sci., 80: 2964-2971.
Direct Link - Bruckental, I., J.D. Oldham and J.D. Sutton, 1980. Glucose and urea kinetics in early lactation. Br. J. Nutr., 44: 33-45.
PubMedDirect Link - Burgos, S.A., J.G. Fadel and E.J. de Peters, 2007. Prediction of ammonia emission from dairy cattle manure based on milk urea nitrogen: Relation of milk urea nitrogen to urine urea nitrogen excretion. J. Dairy Sci., 90: 5499-5508.
CrossRef - Butler, W.R., J.J. Calaman and S.W. Beam, 1996. Plasma and milk urea nitrogen in relation to pregnancy rate in lactating dairy cattle. J. Anim. Sci., 74: 858-865.
CrossRefPubMedDirect Link - Canfield, R.W., C.J. Sniffen and W.R. Butler, 1990. Effects of excess degradable protein on postpartum reproduction and energy balance in dairy cattle. J. Dairy Sci., 73: 2342-2349.
CrossRefDirect Link - Cannas, A., A. Pes, R. Mancuso, B. Vodret and A. Nudda, 1998. Effect of dietary energy and protein concentration on the concentration of milk urea nitrogen in dairy ewes. J. Dairy Sci., 81: 499-508.
Direct Link - Carlsson, J., J. Bergsrtom and B. Pehrson, 1995. Variations with breed, age, season, yield, stage of lactation and herd in the concentration of urea in bulk milk and individual cows milk. Acta. Vet. Scand., 36: 245-254.
PubMedDirect Link - Carlsson, J. and J. Bergstrom, 1994. The diurnal variation of urea in cow`s milk and how milk fat content, storage and preservation affects analysis by a flow injection technique. Acta Vet. Scand., 35: 67-77.
PubMedDirect Link - Carroll, D.J., B.A. Barton, G.W. Anderson and R.D. Smith, 1988. Influence of protein intake and feeding strategy on reproductive performance of dairy cows. J. Dairy Sci., 71: 3470-3481.
PubMedDirect Link - Ciszuk, A.U. and T. Gebregziabher, 1994. Milk urea as an estimate of urine nitrogen of dairy cows and goats. Acta Agric. Scand., 44: 87-95.
CrossRef - Collard, B.L., P.J. Boettcher, J.C.M. Dekkers, D. Petitclerc and L.R. Schaeffer, 2000. Relationships between energy balance and health traits of dairy cattle in early lactation. J. Dairy Sci., 83: 2683-2690.
Direct Link - Cottrill, B., H.J. Biggadike, C. Collins and R.A. Laven, 2002. Relationship between milk urea concentration and the fertility of dairy cows. Vet. Record, 151: 413-416.
Direct Link - De Peters, E.J. and J.P. Cant, 1992. Nutritional factors influencing the nitrogen composition of bovine milk: A review. J. Dairy Sci., 75: 2043-2070.
CrossRefPubMedDirect Link - De Wit, A.A.C., M.L.F. Ceser and T.A.M. Kruip, 2001. Effect of urea during in vitro maturation on nuclear maturation and embryo development of bovine cumulus oocyte complexes. J. Dairy Sci., 84: 1800-1804.
Direct Link - De Peters, E.J. and J.D. Ferguson, 1992. Nonprotein nitrogen and protein distribution in the milk of cows. J. Dairy Sci., 75: 3192-3209.
Direct Link - Edwards, L.J., D.A. Williams and D.K. Gardner, 1998. Intracellular pH of the mouse pre-implantation embryo: Amino acids act as buffers of intracellular pH. Hum. Reprod., 13: 3441-3448.
Direct Link - Elrod, C.C. and W.R. Butler, 1993. Reduction of fertility and alteration of uterine pH in heifers fed excess ruminally degradable protein. J. Anim. Sci., 71: 694-701.
PubMedDirect Link - Elrod, C.C., M. van Amburgh and W.R. Butler, 1993. Alterations of pH in response to increased dietary protein in cattle are unique to the uterus. J. Anim. Sci., 71: 702-706.
Direct Link - Ferguson, J.D. and W. Chalupa, 1989. Impact of protein nutrition on reproduction in dairy cows. J. Dairy Sci., 72: 746-766.
CrossRef - Ferguson, J.D., T. Blanchard, D.T. Galligan, D. Hoshall, and W. Chalupa, 1988. Infertility in dairy cattle fed a high percentage of protein degradable in the rumen. J. Am. Vet. Med. Assoc., 192: 659-662.
PubMedDirect Link - Ferguson, J.D., D.T. Galligan, T. Blanchard and M. Reeves, 1993. Serum urea nitrogen and conception rate: the usefulness of test information. J. Dairy Sci., 76: 3742-3746.
CrossRefPubMedDirect Link - Garcia-Bojalil, C.M., C.R. Staples, C.A. Risco, J.D. Savio and W.W. Thatcher, 1998. Protein degradability and calcium salts of long-chain fatty acids in the diets of lactating dairy cows: Reproductive responses. J. Dairy Sci., 81: 1385-1395.
Direct Link - Garcia-Bojalil, C.M., C.R. Staples, W.W. Thatcher and M. Drost, 1994. Protein intake and development of ovarian follicles and embryos of superovulated non-lactating dairy cows. J. Dairy Sci., 77: 2537-2548.
Direct Link - Gath, V.P., P. Lonergan, M.P. Boland and D. O`Callaghan, 1999. Effect of diet type on establishment of pregnancy and embryo development in heifers. Theriogenology, 51: 224-224.
CrossRef - Godden, S.M., D.F. Kelton, K.D. Lissemore, J.S. Walto and K.E. Leslie et al., 2001. Milk urea testing as a tool to monitor reproductive performance in Ontario dairy herds. J. Dairy Sci., 84: 1397-1406.
Direct Link - Guo, K., E. Russek-Cohen, M.A. Varner and R.A. Kohn, 2004. Effects of milk urea nitrogen and other factors on probability of conception of dairy cows. J. Dairy Sci., 87: 1878-1885.
Direct Link - Gustafsson, A.H. and J. Carlsson, 1993. Effects of silage quality, protein evaluation systems and milk urea content on milk yield and reproduction in dairy cows. Livest. Prod. Sci., 37: 91-105.
CrossRef - Gustafsson, A.H. and D.L. Palmquist, 1993. Diurnal variation of rumen ammonia, serum urea and milk urea in dairy cows at high and low yields. J. Dairy Sci., 76: 475-484.
CrossRefDirect Link - Habib, F.K., S.Q. Maddy and S.R. Stitch, 1980. Zinc induced changes in the progesterone binding properties of the human endometrium. Acta Endocrinol, 94: 99-106.
PubMedDirect Link - Hodgen, G.D. and R.J. Falk, 1971. Estrogen and progesterone regulation of carbonic anhydrase isoenzymes in guinea pig and rabbit uterus. Endocrinology, 89: 859-864.
PubMedDirect Link - Hoeschele, I., 1991. Additive and non-additive genetic variance in female fertility of Holstein. J. Dairy Sci., 74: 1743-1752.
Direct Link - Hojman, D., O. Kroll, G. Adin, M. Gips, B. Hanochi and E. Ezra, 2004. Relationships between milk urea and production, nutrition and fertility traits in israeli dairy herds. J. Dairy Sci., 87: 1001-1011.
Direct Link - Ismail, A., K. Diab and J.K. Hillers, 1996. Effect of selection for milk yield and dietary energy on yield traits, bovine somatotropin and plasma urea nitrogen in dairy cows. J. Dairy Sci., 79: 682-688.
Direct Link - Jaquette, R., A.H. Rakesand W.J. Croom, 1986. Effects of dietary protein on milk, rumen, and blood parameters in dairy cattle fed low fiber diets. J. Dairy Sci., 69: 1026-1034.
Direct Link - Jonker, J.S., R.A. Kohn and R.A. Erdman, 1998. Using milk urea nitrogen to predict nitrogen excretion and utilization efficiency in lactating dairy cows. J. Dairy Sci., 81: 2681-2692.
Direct Link - Jordan, E.R., T.E. Chapman, D.W. Holtan and L.V. Swanson, 1983. Relationship of dietary crude protein to composition of uterine secretions and blood in high producing post partum dairy cows. J. Dairy Sci., 66: 1854-1862.
Direct Link - Jorritsma. R., T. Wensing, T.A.M. Kruip, P.L.A.M. Vos and J.P.T.M. Noordhuizen, 2003. Metabolic changes in early lactation and impaired reproductive performance in dairy cows. Vet. Res., 34: 11-26.
CrossRefDirect Link - Kauffman, A.J. and N.R. St-Pierre, 2001. The relationship of milk urea nitrogen to urine nitrogen excretion in Holstein and Jersey cows. J. Dairy Sci., 84: 2284-2294.
CrossRefDirect Link - Kenny, D.A., M.P. Boland, M.G. Diskin and J.M. Sreenan, 2001. Effect of pasture crude protein and fermentable energy supplementation on blood metabolite and progesterone concentration and embryo survival in heifers. Anim. Sci. Abstr., 73: 501-512.
Direct Link - Klusmeyer, T.H., R.D. McCarthy Jr., J.H. Clark and D.R. Nelson, 1990. Effects of source and amount of protein on ruminal fermentation and passage of nutrients to the small intestine of lactating cows. J. Dairy Sci., 73: 3526-3537.
CrossRefDirect Link - Kohn, R.A., Z. Dou, J.D. Ferguson and R.C. Boston, 1997. A sensitivity analysis of nitrogen losses from dairy farms. J. Environ. Manage., 50: 417-428.
CrossRef - Kohn, R.A., K.F. Kalscheur and E. Russek-Cohen, 2002. Evaluation of models to estimate urinary nitrogen and expected milk urea nitrogen. J. Dairy Sci., 85: 227-233.
Direct Link - Kolver, E.S. and K.L. MacMillan, 1993. Short term changes in selected metabolites in pasture fed dairy cows during peak lactation. Proc. N. Z. Soc. Anim. Prod., 53: 77-81.
Direct Link - Kung, Jr. L. and J.T. Huber, 1983. Performance of high producing cows in early lactation fed protein of varying amounts, sources and degradability. J. Dairy Sci., 66: 227-234.
Direct Link - Leese, H.J., S. Aldridge and K.S. Jeffries, 1979. The movement of amino acids into rabbit oviductal fluid. J. Reprod. Fertil., 56: 623-626.
Direct Link - Mauer, R.R. and H.M. Beier, 1976. Uterine proteins and development in vitro of rabbit preimplantation embryos. J. Reprod. Fertil., 48: 33-41.
CrossRef - McEvoy, T., J. Robinson, R. Aitken, P. Findlay and I. Robertson, 1997. Dietary excesses of urea influence the viability and metabolism of pre-implantation sheep embryos and may affect fetal growth among survivors. Anim. Reprod. Sci., 47: 71-90.
PubMedDirect Link - Miettinen, P.V. and R.O. Juvonen, 1990. Diurnal variation of serum and milk urea levels in dairy cows. Acta Agric. Scand., 40: 289-296.
CrossRef - Ng-Kwai-Hang, K.F., J.F. Hayes, J.E. Moxley and H.G. Monardes, 1985. Percentages of protein and nonprotein nitrogen with varying fat and somatic cells in bovine milk. J. Dairy Sci., 68: 1257-1262.
Direct Link - Ocon, O.M. and P.J. Hansen, 2003. Disruption of bovine oocytes and preimplantation embryos by urea and acidic pH. J. Dairy Sci., 86: 1194-1200.
Direct Link - Oldham, J.D., 1984. Protein-energy interrelationships in dairy cows. J. Dairy Sci., 67: 1090-1114.
CrossRefPubMedDirect Link - Oltner, R. and H. Wiktorsson, 1983. Urea concentrations in milk and blood as influenced by feeding varying amounts of protein and energy to dairy cows. Livestock Prod. Sci., 10: 457-467.
CrossRefDirect Link - Oltner, R., M. Emanuelson and H. Wiktorsson, 1985. Urea concentration in milk in relation to milk yield, live weight, lactation number and amount and composition of feed given to dairy cows. Livest. Prod. Sci., 12: 47-57.
CrossRef - Rajala-Schultz, P.J., W.J.A. Saville, G.S. Frazer and T.E. Wittum, 2001. Association between milk urea nitrogen and fertility in Ohio dairy cows. J. Dairy Sci., 84: 482-489.
Direct Link - Refsdal, A.O., 1983. Urea in bulk milk as compared to the herd mean of urea in blood. Acta Vet. Scand., 24: 518-520.
PubMedDirect Link - Refsdal, A.O., L. Baerve and R. Brutflot, 1985. Urea concentration in bulk milk as an indicator of the protein supply at the herd level. Acta Vet. Scand., 26: 153-163.
PubMedDirect Link - Rhoads, M.L., R.O. Gilbert, M.C. Lucy and W.R. Butler, 2004. Effects of urea infusion on the uterine luminal environment of dairy cows. J. Dairy Sci., 87: 2896-2901.
CrossRefDirect Link - Rhoads, M.L., R.P. Rhoads, R.O. Gilbert, R. Toole and W.R. Butler, 2006. Detrimental effects of high plasma urea nitrogen levels on viability of embryos from lactating dairy cows. Anim. Reprod. Sci., 91: 1-10.
CrossRef - Rodriguez-Martinez, H., E. Ekstedt and Y. Ridderstrale, 1991. Histochemical localization of carbonic anhydrase in the female genitalia of pigs during the oestrous cycle. Acta Anat., 140: 41-47.
CrossRef - Ropstad, E. and A.O. Refsdal, 1987. Herd reproduction performance related to urea concentration in bulk milk. Acta Vet. Scand., 28: 55-63.
PubMedDirect Link - Ropstad, E., L. Vik-Mo, and A.O. Refsdal, 1989. Levels of milk urea, plasma constituents and rumen liquid ammonia in relation to the feeding of dairy cows during early lactation. Acta Vet. Scand., 30: 199-208.
PubMedDirect Link - Roseler, D.K., J.D. Ferguson, C.J. Sniffen and J. Herrema. 1993. Dietary protein degradability effects on plasma and milk urea nitrogen and milk nonprotein nitrogen in Holstein cows. J. Dairy Sci., 76: 525-534.
CrossRefDirect Link - Roy, B., R.K. Mehla and S.K. Sirohi, 2003. Influence of non-nutritional factors on urea and protein concentration in milk of Murrah buffaloes. Asian Aust. J. Anim. Sci., 16: 1285-1290.
Direct Link - Roy, B., R.K. Mehla and S.K. Sirohi, 2005. Influence of dietary feeding regimens on milk urea levels in Murrah buffaloes. Asian Aust. J. Anim. Sci., 18: 973-979.
Direct Link - Schepers, A.J. and R.G.M. Meijer, 1998. Evaluation of the urtiliztion of dietary nitrogen by dairy cows based on urea concentration in milk. J. Dairy Sci., 81: 579-584.
Direct Link - Schrick, F.N., E.K. Inskeep and R.L. Butcher, 1993. Pregnancy rates for embryos transferred from early postpartum beef cows into recipients with normal estrous cycles. Biol. Reprod., 49: 617-621.
Direct Link - Sinclair, K.D., M. Kuran, F.E. Gebbie, R. Webband and T.G. McEvoy, 2000. Nitrogen metabolism and fertility in cattle: II. Development of oocytes recovered from heifers offered diets differing in their rate of nitrogen release in the rumen. J. Anim. Sci., 78: 2670-2680.
PubMed - Sonderman, J.P. and L.L. Larson, 1989. Effect of dietary protein and exogenous gonadotropin-realising hormone on circulating progesterone concentrations and performance of Holstein cows. J. Dairy Sci., 72: 2179-2183.
Direct Link - Staples, C.R., J.M. Burke and W.W. Thatcher, 1998. Influence of supplemental fats of reproductive tissues and performance of lactating cows. J. Dairy Sci., 81: 856-871.
Direct Link - Swanson, L.V., 1989. Interaction of nutrition and reproduction. J. Dairy Sci., 72: 746-766.
Direct Link - Vandehaar, M.J., 1998. Efficiency of nutrient use and relationship to profitability on dairy farms. J. Dairy Sci., 81: 272-282.
PubMedDirect Link - Vestweber, J.G.E., F.K. Al-Ani and D.E. Johnson, 1989. Udder edema in cattle: Effects of diuretics (furosemide, hydrochlorothiazide, acetazolamide, and 50% dextrose) on serum and urine electrolytes. Am. J. Vet. Res., 50: 1323-1328.
PubMedDirect Link - Wittwer, F.G., P. Gallardo, J. Reyes and H. Opitz, 1999. Bulk milk urea concentrations and their relationship with cow fertility in grazing dairy herds in Southern Chile. Prev. Vet. Med., 38: 159-166.
PubMedDirect Link - Butler, W.R., D.J.R. Cherney and C.C. Elrod, 1995. Milk urea nitrogen (MUN) analysis: Field trial results on conception rates and dietary inputs. Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, (CNCFM'95), Rochester, NY., pp: 89-95.
Direct Link - Nelson, A.J., 1996. Practical application of MUN analyses. Proceedings of the American Association of Bovine Practitioners Conference, Sept. 12-14, San Diego, CA., pp: 85-95.
Direct Link