
M. Sadeghi
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran
M. Moradi Shahr-e-Babak
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran
G. Rahimi
Department of Animal Science, Faculty of Agriculture, University of Mazandaran, Sari, Iran
A. Nejati Javaremi
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran
Asian Journal of Animal Sciences
Year: 2008 | Volume: 2 | Issue: 1 | Page No.: 1-6







ABSTRACT
The objective of this study was to obtain genetic marker information in the Iranian Holstein bulls by analyzing bGH gene polymorphism and to verify its association with milk production traits. One sample including 134 Iranian Holstein bulls was genotyped at bGH gene restriction sites (bGH-AluI). Gene frequencies of L and V alleles were 0.936 and 0.064, respectively. Genotypes are distributed according to the Hardy-Wienberg equilibrium. Differences in the milk and fat yield between the two genotypes approached significance (p≤0.10). Bulls with LL genotype had higher milk, fat yield compared to LV genotypes. Present study showed that the L allele was associated with protein yield (p≤0.021). No associations (p>0.1) were found between the genotypes and fat and protein percent. The dominant L allele at the bGH locus favours higher production of milk protein. It can be concluded that this locus is a QTL or is in disequilibrium linkage with one or more tightly linked QTL. Thus it could be a useful DNA marker for milk production traits in dairy cattle breeding.
PDF Abstract XML References Citation
How to cite this article
M. Sadeghi, M. Moradi Shahr-e-Babak, G. Rahimi and A. Nejati Javaremi, 2008. Association Between Gene Polymorphism of Bovine Growth Hormone and Milk Traits in the Iranian Holstein Bulls. Asian Journal of Animal Sciences, 2: 1-6.
DOI: 10.3923/ajas.2008.1.6
URL: https://scialert.net/abstract/?doi=ajas.2008.1.6
DOI: 10.3923/ajas.2008.1.6
URL: https://scialert.net/abstract/?doi=ajas.2008.1.6
INTRODUCTION
By selecting the best performing animals on phenotypic variants (milk, fat, or protein yield) as reproducers, breeders have increased the frequency of favorable alleles influencing the interesting traits. Moreover, in dairy cattle, genetic progress is essentially based on the use of genetically superior sires whose identification requires time (5-6 years) and money (1000, 000 US$) (Parmentier et al., 1999). Furthermore, the exact molecular natures of the target genes remain essentially unknown, particularly for production traits that are often controlled by a pool of genes. At the present time, the two major strategies developed to detect Quantitative Trait Loci (QTL) are the candidate gene approach and the positional genetic approach. The somatotropin axis contains the most promising candidates in this respect, as it strongly regulates milk production. Current knowledge in dairy biology indicates that genetically superior animals differ from lesser animals mainly in their regulation of nutrient utilization and that growth hormone exerts a key control in nutrient use, mammary development, growth milk yield (Bauman, 1999) and also modulates intermediary metabolism and other physiological processes e.g., aging and immune responsiveness. Thus the Growth Hormone (GH) gene is a promising candidate gene worth studying for its effects on milk-related traits. GH is a part of a multiple-gene family that contains prolactin and the placental lactogens and has been mapped by in situ hybridization to bovine chromosome 19 (Zwierzchowski et al., 2002) bovine Growth Hormone (bGH) is a 22 kDa pituitary hormone composed of 191 amino acid residues.
Selection for milk yield has been shown to be associated with increased blood levels of GH (Bauman, 1999). GH is synthesized in the pituitary gland and coded for by a single gene consisting of five introne and four exon. Several base substitution polymorphisms have been detected both in the promoter and the coding and none coding regions. One of the base substitutions gives rise to an amino acid change at position 127 in the peptide, where a Leusine is exchanged for Valine. Identification of such mutations would permit selection at the DNA level without necessitating measurement of GH levels.
Danish Jersey calves with Leu/Leu genotype had a higher PEAK than calves with the Val/Val genotype, whereas the Leu/Val genotype had an intermediate response. PEAK was the mean GH of blood sampled at 10, 15 and 20 min following inducement with GHRH (Sorensen et al., 2002).
Grochowska et al. (2001) reported that differences between the Leu/Leu and the Val/Val genotypes for carcass gain and weight of meat in the carcass were significant (p≤0.05). Moreover, differences in the size of the GH peak between the two homozygotes approached significance (p≤0.10). Schlee et al. (1994) reported significant effects for the LV genotype for carcass gain and meat value but reported no effects for milk breeding values. Associations between Leu/Val polymorphism and milk production traits of cows were found in first lactation (Dybus, 2002). Cows with LL genotype had higher milk, fat and protein yield compared to LV individuals (p≤0.01).
Objectives of this study were to detect genetic variants in bGH gene as candidate gene in Holstein proven bulls and to test the effect of the genetic polymorphisms on milk production traits and also to estimates the average effect of allele substitution.
MATERIALS AND METHODS
Semen samples from 134 Holstein proven bulls were obtained from Animal Breeding Center of Iran. Genomic DNA was extracted according to Zadworney and Kuhnlein (1990).
DNA Amplification With PCR-RFLP
bGH genotypes were identified according to Sorensen et al. (2002). The 282 bp fragment of intron 4 and exon 5 of the bGH gene was amplified using fallowing primers: 5` GTG GGC TTG GGG AGA CAG AT 3` (position 1940) and 5` GTC GTC ACT GCG CAT GTT TG 3` (position 2202).
A single reaction (25 μL) contained the fallowing constituents: 10X PCR buffer (16 mM (NH4)2SO4, 67 mM Tris HCl pH 8.8, 0.1% Tween-20), 2.5 mM MgCl2, 200 μM dNTP, 5 pmoles of each primers, 1 unit Taq polymerase (Metabion) and 50-100 ng of genomic DNA. The amplification program consisted of an initial denaturation at 94°C for 2 min, then 30 cycles of 94°C for 45 sec, 62°C for 60 sec and 72°C for 60 sec and a final extension of 72°C was maintained for 3 min.
A single digestion reaction consisted of 10 μL of PCR product, 4 units (0.4 μL) of AluI enzyme (Fermentas), 1.5 μL Tango buffer and 3.15 μL nuclease-free water. The final reaction volume of 15 μL was incubated at 37°C for 12 h. The fragments were separated on a 3% agarose gel by electrophoresis.
Statistical Analysis
Calculation of gene frequency was based on direct gene count method by:
| (2nLL+nLV)/2n |
and standard error of frequency was calculated as:
| |
| Where: |
| n | = | The sample size |
| p | = | The frequency of L allele |
| nLL and nLV | = | No. of LL and LV types, respectively |
The Χ2-test was used to determine Hardy-Weinberg equilibrium (SAS, 2002).
Breeding values for milk related traits (milk, fat and protein yield; and fat and protein percent) were estimated with the Best Linear Unbiased Procedure (BLUP) based on an animal model with a relationship matrix. The analysis was conducted using AI-REML procedures as programmed in MATVEC software (Wang et al., 2002). The model included animal effect as random effect and age of calving as covariate factor and fixed effect of herd-year-season.
The effect of bGH genotypes on the estimated breeding values for milk related traits was analyzed using least square method of GLM procedure of SAS (2002) software. As the breeding values are the best available estimates of the additive genotype of the bulls, no environmental effects were included in the model. The used model was as fallows:
Yij = μ+Gi+eij |
| Where: |
| yij | = | The breeding value for milk related traits |
| μ | = | The least square means of the traits |
| Gi | = | The effect of the ith genotype (I = 1, 2, 3) |
| eij | = | The random residual effect |
Type III sum of squares were used to evaluate the effect of bGH polymorphism.
Regression analyses were performed in which EBV for milk, fat, protein yield; and fat and protein percent were the dependent variables and the genotype was the independent variable. Average effect of allele substitution was determined by coding genotypes as 0(VV), 1(LV) and 2(LL) to represent the number of L alleles present for the bGH polymorphism. As described by Falconer and Mackay (1996), the regression coefficient (α) estimates the average effect of allele substitution, or the average effect of replacing a V allele with an L allele.
RESULTS AND DISCUSSION
The PCR amplified a 282 bp fragment from intron 4 and exon 5 of the bGH gene. The resulting digestion with AluI enzyme was two alleles. The leucine (L) allele had fragment sizes of 150, 82 and 50 bp, whereas the valine (V) allele had fragments of 150 and 132 bp (Fig. 1). Gene frequencies of L and V alleles were 0.936 and 0.064, respectively.
Genotypes are distributed according to the Hardy-Weinberg equilibrium. The L allele frequency obtained in our study is agreement with those reported by the other researchers, for example, Danish Holstein, 0.93; Danish Red, 0.85; Danish Jersey, 0.51 (Sorensen et al., 2002), Holstein cows, 0.863 (Lee et al., 1996), Polish Friesian bull, 0.64 (Grochowska et al., 2001), Holstein bulls, 0.91 (Yao et al., 1996), Brown Swiss, 1.00; Guernsey, 0.92; Ayshire, 0.72; Jersey, 0.56 (Lucy et al., 1993), Aberdeen Angus, 0.77, Simmental, 0.82 (Luciana et al., 2003). Holstein, Brown Swiss and Guernsey breeds had higher frequencies of the L allele of bGH whereas highest frequencies for the V allele were found in Jersey and Airshire cows. Variants of bGH at position 127 are the result of a point mutation (from CTG to GTG) in the nucleotide sequence. This change converts the leucine codon to the valine codon (Lucy et al., 1993).
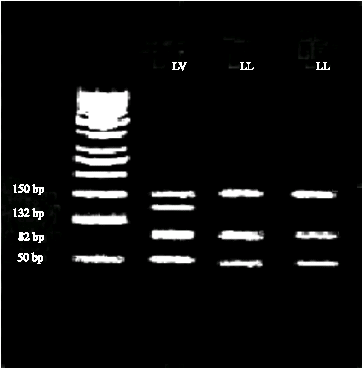 | |
| Fig. 1: | A 3% agarose gel displaying an AluI restriction digest on an amplified portion of bGH. Lane 4 is 50 bp ladder (Fermentase) |
| Table 1: | Least square means of breeding values for milk yield and fat and protein yield and percent in Iranian Holstein bulls with two bGH genotypes |
 | |
| Values are means ± SE | |
The effect of the genotypes of AluI polymorphism on the breeding values for milk related traits was examined using least square methods. Least square means of the two bGH genotypes are shown in Table 1.
Differences in the milk and fat yield between the two genotypes approached significance (p≤0.10). Bulls with LL genotype had higher milk, fat yield compared to LV genotypes. Present study showed that the L allele was associated with protein yield (p≤0.021). No associations (p>0.1) were found between the genotypes and fat and protein percent. Amino acid position 127 is located in the third α-helix of the bGH molecule and is close to residues that are involved in receptor binding. Therefore, changes in the bGH molecule at this location may change interactions of bGH with its receptor and affect growth or milk production.
These results for the GH RFLP are consistent with the results of (Lee et al., 1996; Lucy et al., 1993) who reported that in American Holstein-Friesian cows, the V allele is associated with low milk production. Differences were found between the LL and the VV genotypes for carcass gain and weight of meat in the carcass (Grochowska et al., 2001).
Shariflou et al. (2000) reported that the L allele of GH favoured higher production of milk, fat and protein yield and no significant differences were found between genotypes with percentages of fat and protein.
| Table 2: | Average additive gene substitution effects of alleles of AluI-GH polymorphisms on milk, fat and protein yield; and fat and protein percent |
 | |
Schlee et al. (1994) also reported that in AI bulls of three cattle breeds in Germany, the LL genotype is associated with higher concentrations of circulating growth hormone in different physiological conditions. Yao et al. (1996) reported that there were no significant differences between genotypes LL and LV in Canadian Holstein bulls for milk yield, fat percent and protein yield.
The average effects of replacing a V allele with an L allele are shown in Table 2. The average effects of the gene substitution for AluI were amounted to 177 kg for milk yield, 3.8 kg for fat yield and 5.4 kg for protein yield and -0.04 and 0.03 for fat and protein percent, respectively.
The dominant L allele at the GH locus favours higher production of milk protein. It can be concluded that this locus is a QTL or is in disequilibrium linkage with one or more tightly linked QTL. Either way, it could be a useful DNA marker for milk production traits in dairy cattle breeding.
ACKNOWLEDGMENTS
This research was financially supported by University of Tehran, Iran. Data and pedigree records and semen of Iranian Holstein bulls was provided through the Animal Breeding Center of Iran.
REFERENCES
- Bauman, D.E., 1999. Bovine somatotropin and lactation: From basic science to commercial application. Domest. Animal Endocrinol., 17: 101-116.
Direct Link - Dybus, A., 2002. Associations between Leu/Val polymorphism of growth hormone gene and milk production traits in black-and-white cattle. Archiv Tierzucht, 45: 421-428.
Direct Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Grochowska, R., A. Lunden, L. Zwierzchowski, M. Snochowski and J. Oprzadek, 2001. Association between gene polymorphism of growth hormone and carcass traits in dairy bulls. Animal Sci., 72: 441-447.
CrossRefDirect Link - Luciana, P.M.K.V., D.T. Talhari, A.P. Pereira, L.L. Coutinno and L.C.A. Regitano, 2003. Genetic characterization of Aberdeen Angus cattle using molecular markers. Genet. Mol. Biol., 26: 133-137.
Direct Link - Lucy, M.C., S.D. Hauser, P.J. Eppard, G.G. Krivi, J.H. Clark, D.E. Bauman and R.J. Collier, 1993. Variants of somatotropine in cattle: Gene frequencies in major dairy breeds and associated milk production. Domest. Animal Endocrinol., 10: 325-333.
PubMed - Parmentier, I., D. Portetelle, N. Gengler, A. Pradi and C. Bertozzi et al., 1999. Candidate gene markers associated with somatotropic axis and milk selection. Domestic Anim. Endocrinol., 17: 139-148.
PubMedDirect Link - Schlee, P., R. Graml, E. Schallenberger, D. Schams, O. Rottmann, A. Olbrichaludau and F. Pirchne, 1994. Growth hormone and insulin-like growth factor I concentrations in bulls of various growth hormone genotypes. Theor. Applied Genet., 88: 497-500.
CrossRefDirect Link - Shariflou, M.R., C. Moran and F.W. Nicholas, 2000. Association of the Leu (127) variant of the bovine growth hormone gen with increased yield of milk, fat and protein in Australian Holstein-Friesien. Aust. J. Agric. Res., 51: 515-522.
Direct Link - Sorensen, P., R. Grochowska, L. Holm, M. Henryon and P. Lovendahl, 2002. Polymorphism in the bovine growth hormone gene affects endocrine release in dairy calves. J. Dairy Sci., 85: 1887-1893.
CrossRefDirect Link - Yao, J., S.E. Aggrey, D. Zadworney, J.F. Hayes and U. Kuhnlein, 1996. Sequence variations in the bovine growth hormone gene characterized by single-strand conformation polymorphism (SSCP) analysis and their association with milk production traits in Holsteins. Genetics, 144: 1809-1816.
PubMedDirect Link - Zadworney, D. and U. Kuhnlein, 1990. The identification of the kappa casein genotype in Holstein dairy cattle using polymerase chain reaction. Theor. Applied Genet., 80: 631-634.
CrossRefDirect Link - Zwierzchowski, L., J. Krzyzewski, N. Strzalkowska, E. Siadkowska and Z. Ryniewicz, 2002. Effects of polymorphism of growth hormone, pit-1 and leptin genes, cow's age, lactation stage and somatic cell count on milk yield and composition of Polish black and white cows. Anim. Sci. Pap. Rep., 20: 213-227.